Department of Pediatrics, Baylor College of Medicine, Houston, Texas, United States of America.
PLoS Genet. 2011 Aug;7(8):e1002233. doi: 10.1371/journal.pgen.1002233. Epub 2011 Aug 11.
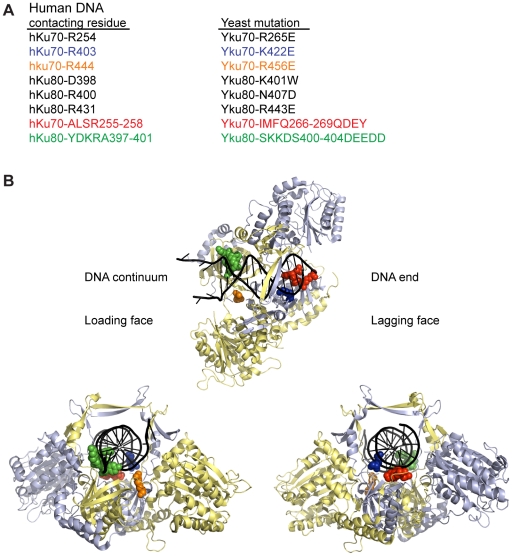
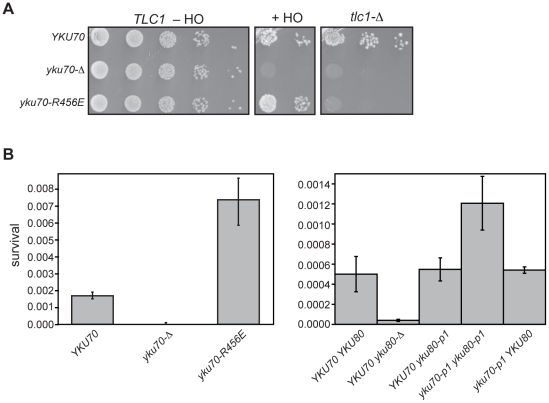
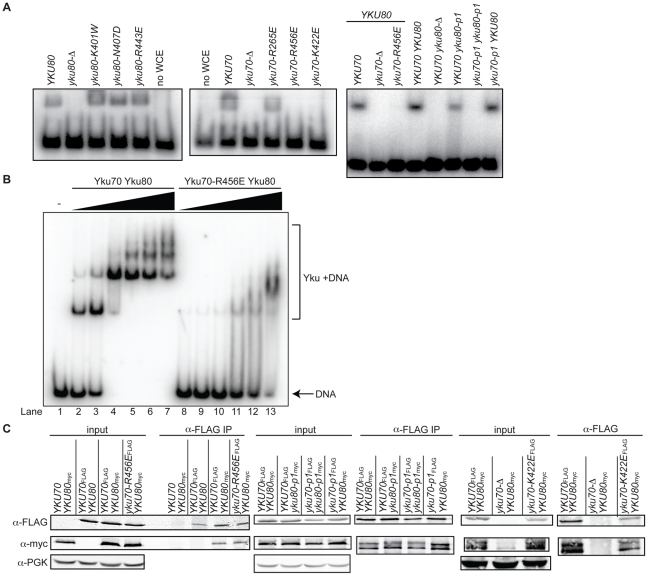
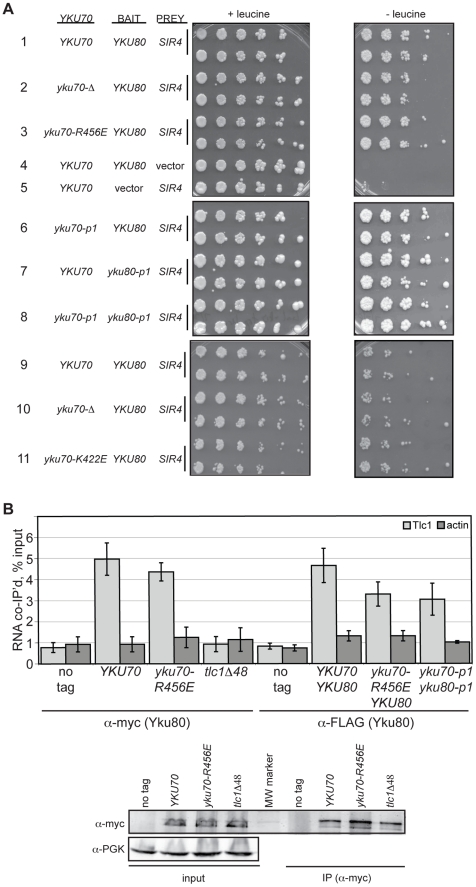
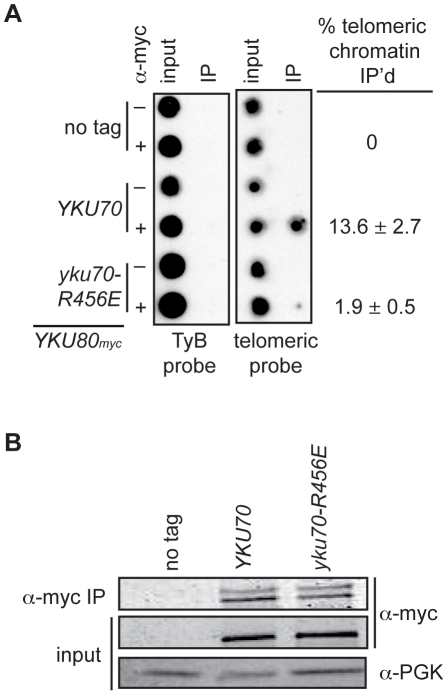
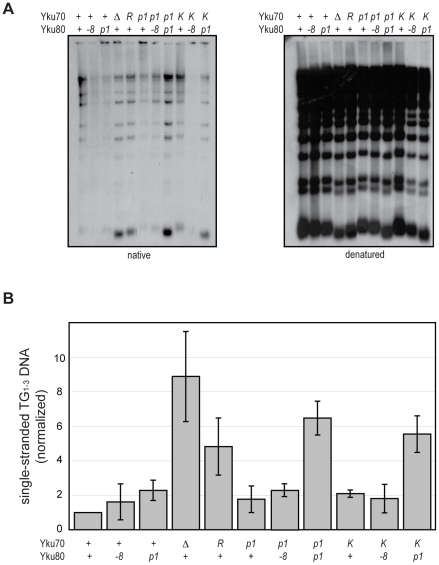
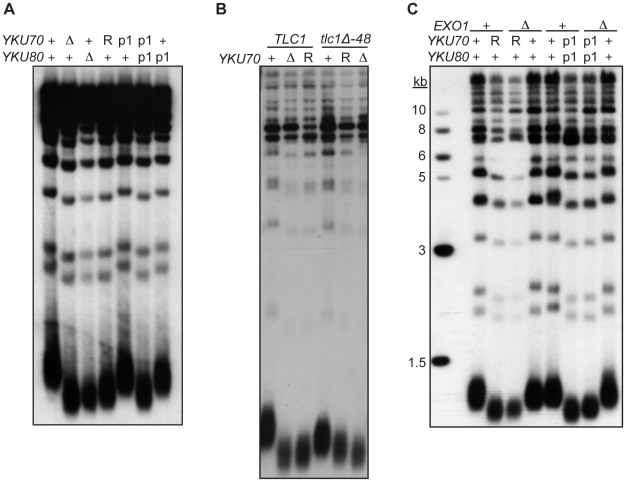
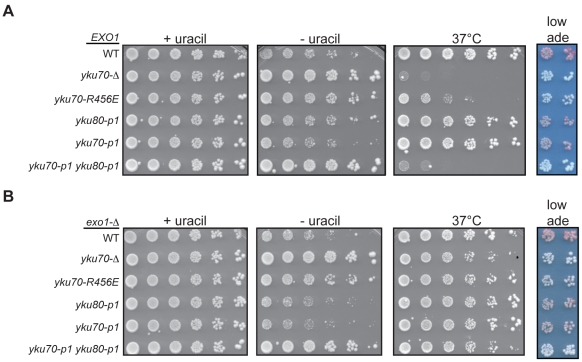
The Ku heterodimer associates with the Saccharomyces cerevisiae telomere, where it impacts several aspects of telomere structure and function. Although Ku avidly binds DNA ends via a preformed channel, its ability to associate with telomeres via this mechanism could be challenged by factors known to bind directly to the chromosome terminus. This has led to uncertainty as to whether Ku itself binds directly to telomeric ends and whether end association is crucial for Ku's telomeric functions. To address these questions, we constructed DNA end binding-defective Ku heterodimers by altering amino acid residues in Ku70 and Ku80 that were predicted to contact DNA. These mutants continued to associate with their known telomere-related partners, such as Sir4, a factor required for telomeric silencing, and TLC1, the RNA component of telomerase. Despite these interactions, we found that the Ku mutants had markedly reduced association with telomeric chromatin and null-like deficiencies for telomere end protection, length regulation, and silencing functions. In contrast to Ku null strains, the DNA end binding defective Ku mutants resulted in increased, rather than markedly decreased, imprecise end-joining proficiency at an induced double-strand break. This result further supports that it was the specific loss of Ku's telomere end binding that resulted in telomeric defects rather than global loss of Ku's functions. The extensive telomere defects observed in these mutants lead us to propose that Ku is an integral component of the terminal telomeric cap, where it promotes a specific architecture that is central to telomere function and maintenance.
Ku 异源二聚体与酿酒酵母端粒结合,影响端粒结构和功能的几个方面。尽管 Ku 通过预先形成的通道强烈结合 DNA 末端,但通过这种机制与端粒结合的能力可能会受到直接与染色体末端结合的因素的挑战。这导致了不确定性,即 Ku 本身是否直接结合端粒末端,以及末端结合是否对 Ku 的端粒功能至关重要。为了解决这些问题,我们通过改变预测与 DNA 接触的 Ku70 和 Ku80 中的氨基酸残基来构建 DNA 末端结合缺陷型 Ku 异源二聚体。这些突变体继续与它们已知的与端粒相关的伴侣结合,例如 Sir4,这是端粒沉默所必需的因子,以及 TLC1,端粒酶的 RNA 成分。尽管存在这些相互作用,但我们发现 Ku 突变体与端粒染色质的结合明显减少,并且对端粒末端保护、长度调节和沉默功能具有类似缺失的缺陷。与 Ku 缺失菌株相反,DNA 末端结合缺陷型 Ku 突变体导致在诱导的双链断裂处非精确末端连接效率显著增加,而不是明显降低。这一结果进一步支持了正是 Ku 对端粒末端结合的特异性丧失导致了端粒缺陷,而不是 Ku 功能的全局丧失。这些突变体中观察到的广泛端粒缺陷导致我们提出 Ku 是末端端粒帽的一个组成部分,它促进了一种特定的结构,这是端粒功能和维持的核心。