Laboratory of Molecular Biology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland, USA.
mBio. 2011 Nov 22;2(6). doi: 10.1128/mBio.00257-11. Print 2011.
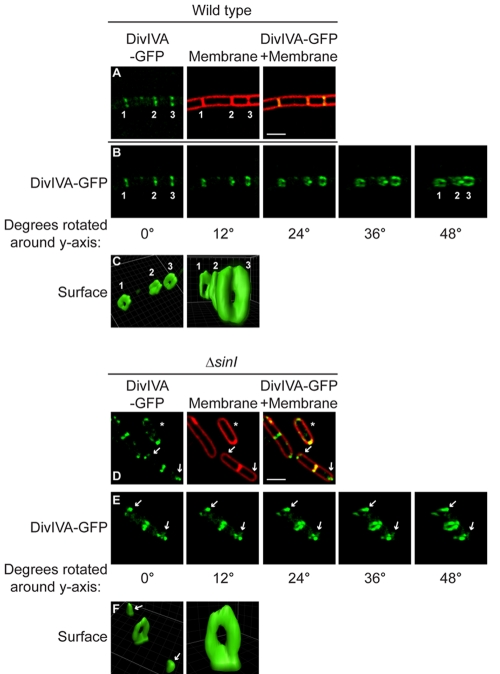
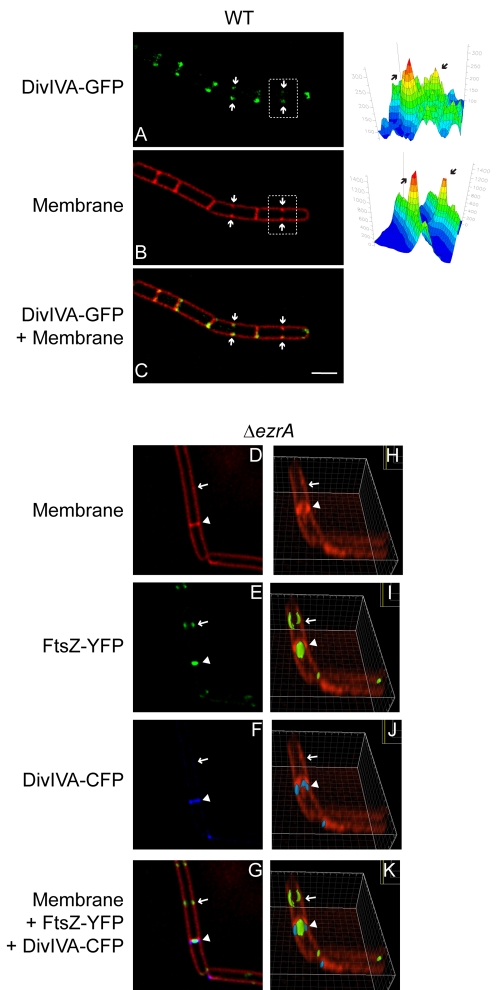
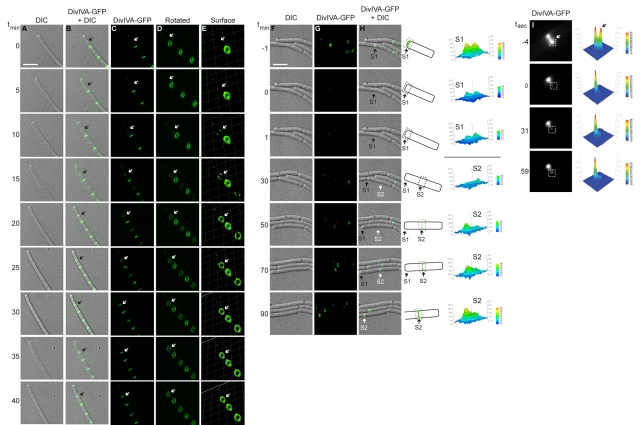
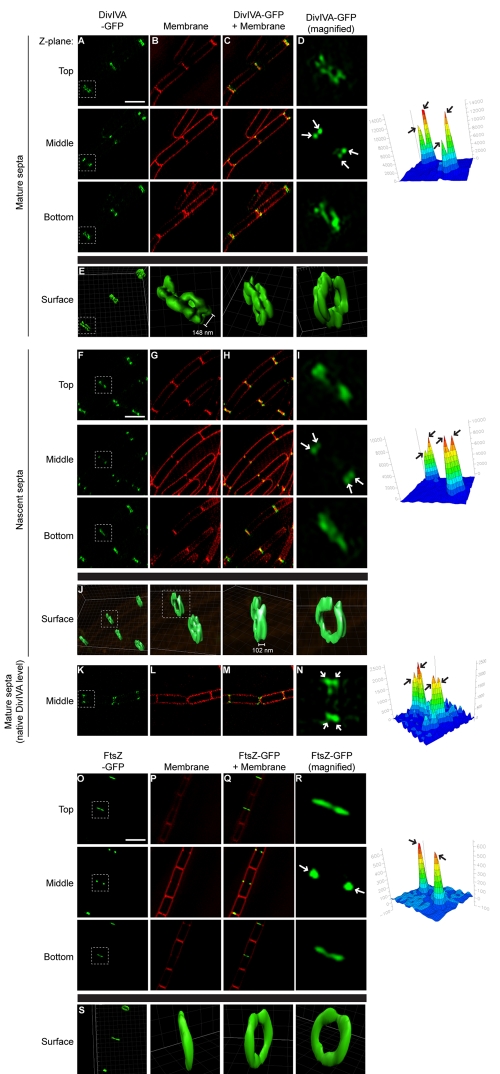
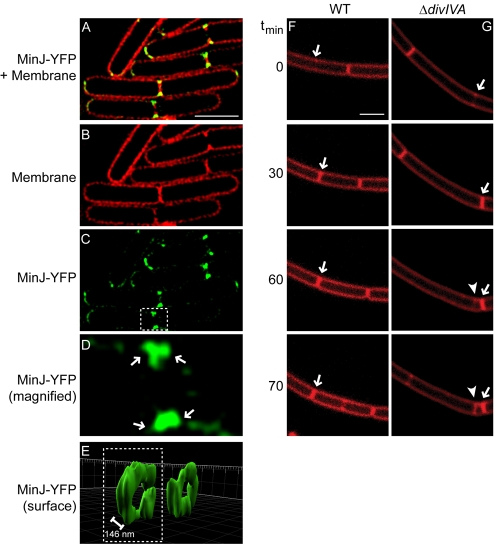
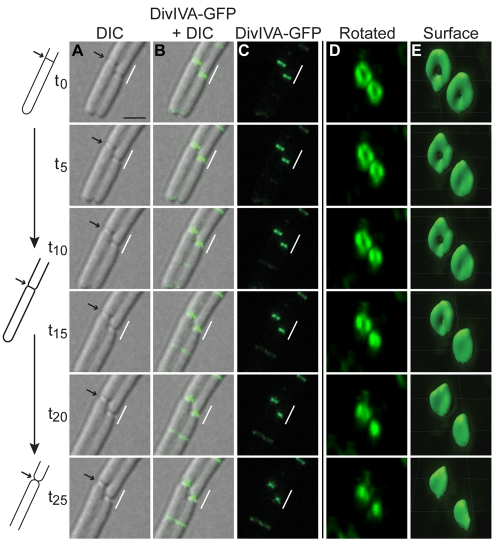
The assembly of the cell division machinery at midcell is a critical step of cytokinesis. Many rod-shaped bacteria position septa using nucleoid occlusion, which prevents division over the chromosome, and the Min system, which prevents division near the poles. Here we examined the in vivo assembly of the Bacillus subtilis MinCD targeting proteins DivIVA, a peripheral membrane protein that preferentially localizes to negatively curved membranes and resembles eukaryotic tropomyosins, and MinJ, which recruits MinCD to DivIVA. We used structured illumination microscopy to demonstrate that both DivIVA and MinJ localize as double rings that flank the septum and first appear early in septal biosynthesis. The subsequent recruitment of MinCD to these double rings would separate the Min proteins from their target, FtsZ, spatially regulating Min activity and allowing continued cell division. Curvature-based localization would also provide temporal regulation, since DivIVA and the Min proteins would localize to midcell after the onset of division. We use time-lapse microscopy and fluorescence recovery after photobleaching to demonstrate that DivIVA rings are highly stable and are constructed from newly synthesized DivIVA molecules. After cell division, DivIVA rings appear to collapse into patches at the rounded cell poles of separated cells, with little or no incorporation of newly synthesized subunits. Thus, changes in cell architecture mediate both the initial recruitment of DivIVA to sites of cell division and the subsequent collapse of these rings into patches (or rings of smaller diameter), while curvature-based localization of DivIVA spatially and temporally regulates Min activity.
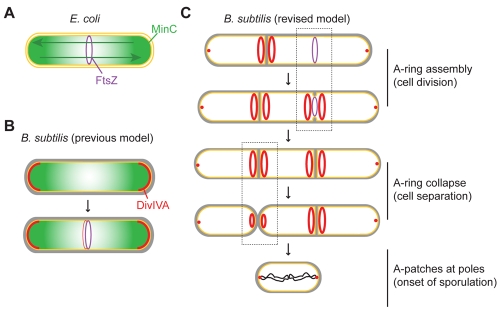
The Min systems of Escherichia coli and Bacillus subtilis both inhibit FtsZ assembly, but one key difference between these two species is that whereas the E. coli Min proteins localize to the poles, the B. subtilis proteins localize to nascent division sites by interaction with DivIVA and MinJ. It is unclear how MinC activity at midcell is regulated to prevent it from interfering with FtsZ engaged in medial cell division. We used superresolution microscopy to demonstrate that DivIVA and MinJ, which localize MinCD, assemble double rings that flank active division sites and septa. This curvature-based localization mechanism holds MinCD away from the FtsZ ring at midcell, and we propose that this spatial organization is the primary mechanism by which MinC activity is regulated to allow division at midcell. Curvature-based localization also conveys temporal regulation, since it ensures that MinC localizes after the onset of division.
细胞分裂装置在细胞中部的组装是细胞分裂的关键步骤。许多杆状细菌使用类核阻塞来定位隔膜,类核阻塞阻止了染色体的分裂,而 Min 系统则防止了靠近极的分裂。在这里,我们研究了枯草芽孢杆菌 MinCD 靶向蛋白 DivIVA 的体内组装,DivIVA 是一种优先定位于负曲率膜的外周膜蛋白,类似于真核细胞的原肌球蛋白,以及 MinJ,它将 MinCD 招募到 DivIVA。我们使用结构光照明显微镜证明 DivIVA 和 MinJ 都定位于隔膜两侧的双环中,并且这些双环最早出现在隔膜生物合成的早期。随后 MinCD 对这些双环的募集将 Min 蛋白与其靶标 FtsZ 分离,从而在空间上调节 Min 活性并允许细胞继续分裂。基于曲率的定位还提供了时间调节,因为 DivIVA 和 Min 蛋白在细胞分裂开始后才会定位于细胞中部。我们使用延时显微镜和光漂白后荧光恢复来证明 DivIVA 环非常稳定,并且由新合成的 DivIVA 分子构建。细胞分裂后,DivIVA 环似乎在分离细胞的圆形细胞极处崩溃成斑块,很少或没有新合成的亚基掺入。因此,细胞结构的变化介导了 DivIVA 最初被招募到细胞分裂部位,以及随后这些环崩溃成斑块(或直径较小的环),而 DivIVA 的基于曲率的定位在空间和时间上调节 Min 活性。
大肠杆菌和枯草芽孢杆菌的 Min 系统都抑制 FtsZ 组装,但这两个物种之间的一个关键区别是,大肠杆菌的 Min 蛋白定位于极,而枯草芽孢杆菌的蛋白通过与 DivIVA 和 MinJ 相互作用定位于新的分裂部位。MinC 在细胞中部的活性如何被调节以防止其干扰参与中部分裂的 FtsZ 尚不清楚。我们使用超分辨率显微镜证明,定位 MinCD 的 DivIVA 和 MinJ 组装了定位于活跃分裂部位和隔膜两侧的双环。这种基于曲率的定位机制将 MinCD 与 FtsZ 环在细胞中部隔开,我们提出这种空间组织是调节 MinC 活性以允许在细胞中部进行分裂的主要机制。基于曲率的定位也传递了时间调节,因为它确保 MinC 在细胞分裂开始后定位。