Ontario Cancer Institute and Department of Medical Biophysics, University of Toronto, Ontario M5G 1L7, Canada.
Nature. 2012 Jan 29;483(7387):108-12. doi: 10.1038/nature10751.
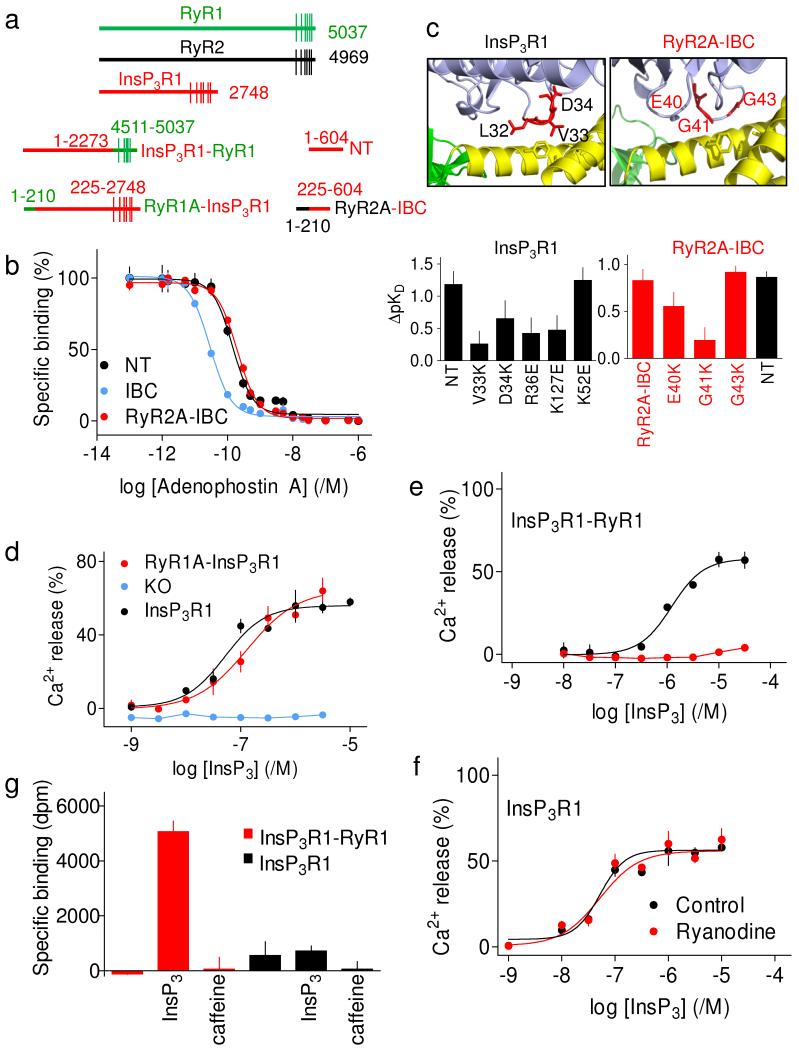
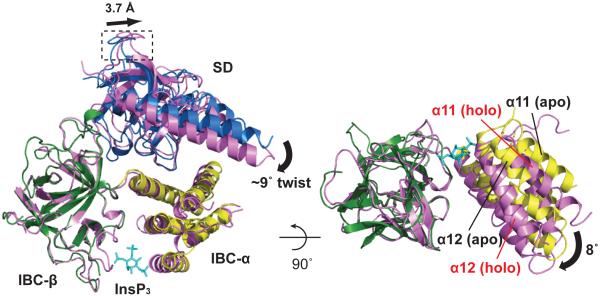
Inositol-1,4,5-trisphosphate receptors (InsP(3)Rs) and ryanodine receptors (RyRs) are tetrameric intracellular Ca(2+) channels. In each of these receptor families, the pore, which is formed by carboxy-terminal transmembrane domains, is regulated by signals that are detected by large cytosolic structures. InsP(3)R gating is initiated by InsP(3) binding to the InsP(3)-binding core (IBC, residues 224-604 of InsP(3)R1) and it requires the suppressor domain (SD, residues 1-223 of InsP(3)R1). Here we present structures of the amino-terminal region (NT, residues 1-604) of rat InsP(3)R1 with (3.6 Å) and without (3.0 Å) InsP(3) bound. The arrangement of the three NT domains, SD, IBC-β and IBC-α, identifies two discrete interfaces (α and β) between the IBC and SD. Similar interfaces occur between equivalent domains (A, B and C) in RyR1 (ref. 9). The orientations of the three domains when docked into a tetrameric structure of InsP(3)R and of the ABC domains docked into RyR are remarkably similar. The importance of the α-interface for activation of InsP(3)R and RyR is confirmed by mutagenesis and, for RyR, by disease-causing mutations. Binding of InsP(3) causes partial closure of the clam-like IBC, disrupting the β-interface and pulling the SD towards the IBC. This reorients an exposed SD loop ('hotspot' (HS) loop) that is essential for InsP(3)R activation. The loop is conserved in RyR and includes mutations that are associated with malignant hyperthermia and central core disease. The HS loop interacts with an adjacent NT, suggesting that activation re-arranges inter-subunit interactions. The A domain of RyR functionally replaced the SD in full-length InsP(3)R, and an InsP(3)R in which its C-terminal transmembrane region was replaced by that from RyR1 was gated by InsP(3) and blocked by ryanodine. Activation mechanisms are conserved between InsP(3)R and RyR. Allosteric modulation of two similar domain interfaces within an N-terminal subunit reorients the first domain (SD or A domain), allowing it, through interactions of the second domain of an adjacent subunit (IBC-β or B domain), to gate the pore.
肌醇 1,4,5-三磷酸受体(InsP(3)Rs)和兰尼碱受体(RyRs)是四聚体细胞内 Ca(2+)通道。在这两个受体家族中,由羧基末端跨膜结构域形成的孔由通过大胞质结构检测到的信号调节。InsP(3)R 的门控由 InsP(3)与 InsP(3)结合核心(IBC,InsP(3)R1 的残基 224-604)结合引发,并且需要抑制域(SD,InsP(3)R1 的残基 1-223)。在这里,我们展示了与(3.6 Å)和不与(3.0 Å)InsP(3)结合的大鼠 InsP(3)R1 的氨基末端结构域(NT,残基 1-604)的结构。三个 NT 结构域(SD、IBC-β 和 IBC-α)的排列确定了 IBC 和 SD 之间的两个离散界面(α 和 β)。在 RyR1 中的等效结构域(A、B 和 C)之间也存在类似的界面(ref.9)。当将三个结构域对接入 InsP(3)R 的四聚体结构中以及将 ABC 结构域对接入 RyR 时,它们的取向非常相似。对 InsP(3)R 和 RyR 的激活重要性的α-界面通过突变得到证实,并且对于 RyR ,通过引起疾病的突变得到证实。InsP(3)的结合导致蛤壳样 IBC 的部分关闭,破坏β-界面并将 SD 拉向 IBC。这使暴露的 SD 环(“热点”(HS)环)重新定向,这对于 InsP(3)R 的激活至关重要。该环在 RyR 中保守,包括与恶性高热和中央核心疾病相关的突变。HS 环与相邻的 NT 相互作用,表明激活重新排列了亚基间的相互作用。RyR 的 A 结构域完全替代了全长 InsP(3)R 的 SD,并且其 C 末端跨膜结构域被 RyR1 取代的 InsP(3)R 可被 InsP(3)门控并被兰尼碱阻断。InsP(3)R 和 RyR 之间的激活机制是保守的。在 N 端亚基内的两个类似的结构域界面的变构调节重新定向第一个结构域(SD 或 A 结构域),从而允许它通过与相邻亚基的第二个结构域(IBC-β 或 B 结构域)的相互作用,使孔门控。