Theoretical Molecular Biophysics Group, Max-Planck-Institute for Biophysical Chemistry, Göttingen, Germany.
PLoS One. 2012;7(6):e38302. doi: 10.1371/journal.pone.0038302. Epub 2012 Jun 28.
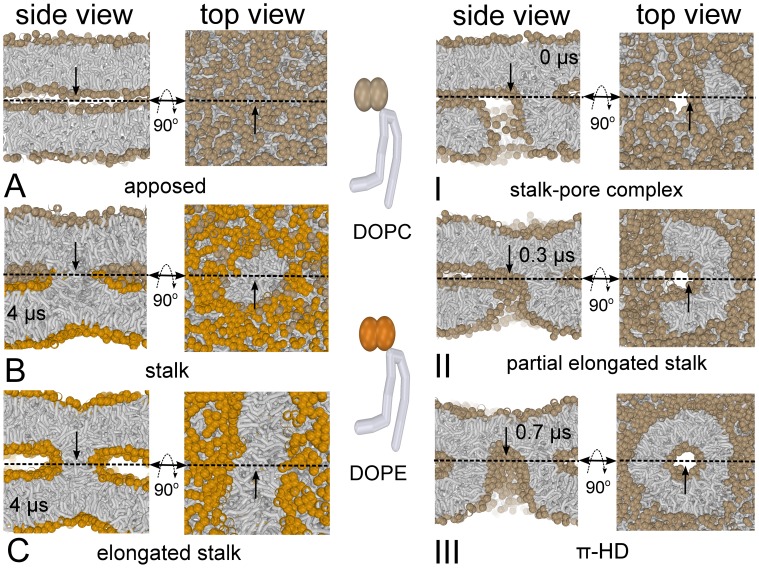
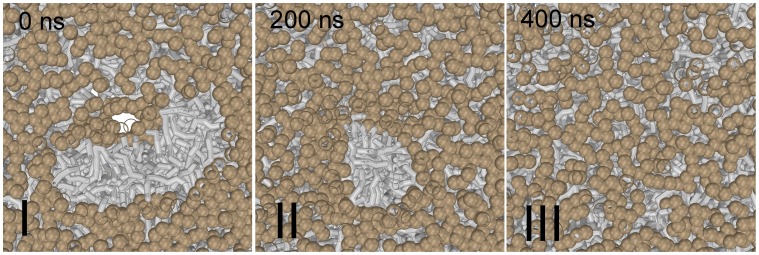
Our molecular simulations reveal that wild-type influenza fusion peptides are able to stabilize a highly fusogenic pre-fusion structure, i.e. a peptide bundle formed by four or more trans-membrane arranged fusion peptides. We rationalize that the lipid rim around such bundle has a non-vanishing rim energy (line-tension), which is essential to (i) stabilize the initial contact point between the fusing bilayers, i.e. the stalk, and (ii) drive its subsequent evolution. Such line-tension controlled fusion event does not proceed along the hypothesized standard stalk-hemifusion pathway. In modeled influenza fusion, single point mutations in the influenza fusion peptide either completely inhibit fusion (mutants G1V and W14A) or, intriguingly, specifically arrest fusion at a hemifusion state (mutant G1S). Our simulations demonstrate that, within a line-tension controlled fusion mechanism, these known point mutations either completely inhibit fusion by impairing the peptide's ability to stabilize the required peptide bundle (G1V and W14A) or stabilize a persistent bundle that leads to a kinetically trapped hemifusion state (G1S). In addition, our results further suggest that the recently discovered leaky fusion mutant G13A, which is known to facilitate a pronounced leakage of the target membrane prior to lipid mixing, reduces the membrane integrity by forming a 'super' bundle. Our simulations offer a new interpretation for a number of experimentally observed features of the fusion reaction mediated by the prototypical fusion protein, influenza hemagglutinin, and might bring new insights into mechanisms of other viral fusion reactions.
我们的分子模拟揭示,野生型流感融合肽能够稳定一种高度融合的预融合结构,即由四个或更多跨膜排列的融合肽形成的肽束。我们推断,这种肽束周围的脂质边缘具有非零的边缘能(线张力),这对于(i)稳定融合双层之间的初始接触点,即柄部,以及(ii)驱动其随后的演变至关重要。这种线张力控制的融合事件不会沿着假设的标准柄部-半融合途径进行。在模拟的流感融合中,流感融合肽中的单点突变要么完全抑制融合(突变体 G1V 和 W14A),要么有趣的是,专门将融合固定在半融合状态(突变体 G1S)。我们的模拟表明,在线张力控制的融合机制中,这些已知的点突变要么通过损害肽稳定所需肽束的能力完全抑制融合(G1V 和 W14A),要么稳定导致动力学捕获的半融合状态的持久束(G1S)。此外,我们的结果进一步表明,最近发现的渗漏融合突变体 G13A,已知在脂质混合之前促进靶膜的明显渗漏,通过形成“超级”束降低膜的完整性。我们的模拟为原型融合蛋白——流感血凝素介导的融合反应的许多实验观察到的特征提供了新的解释,并可能为其他病毒融合反应的机制带来新的见解。