Department of Biochemistry, School of Life Sciences, University of Hyderabad, Hyderabad, Andhra Pradesh, India.
PLoS One. 2012;7(7):e41925. doi: 10.1371/journal.pone.0041925. Epub 2012 Jul 30.
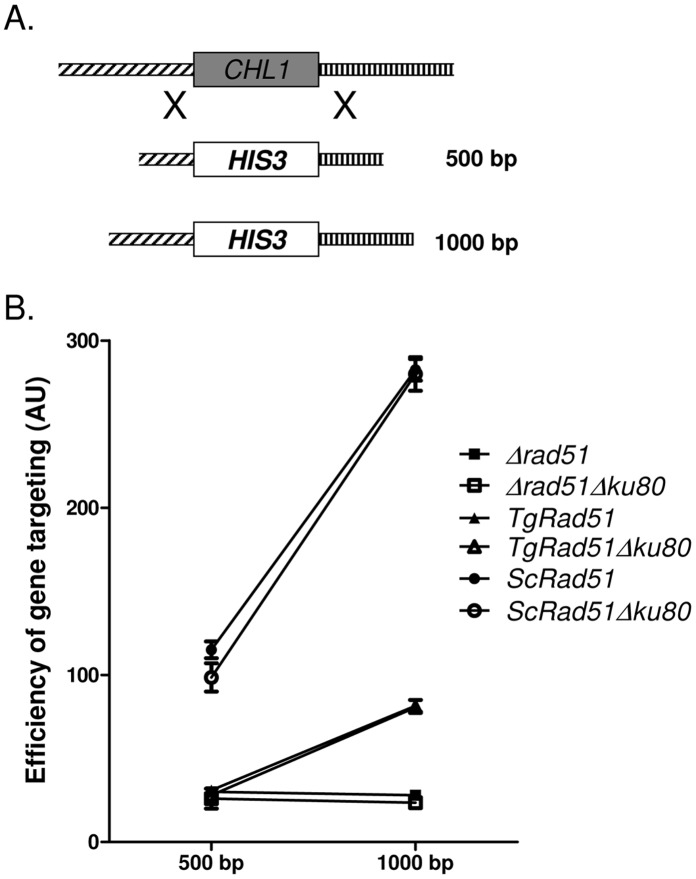
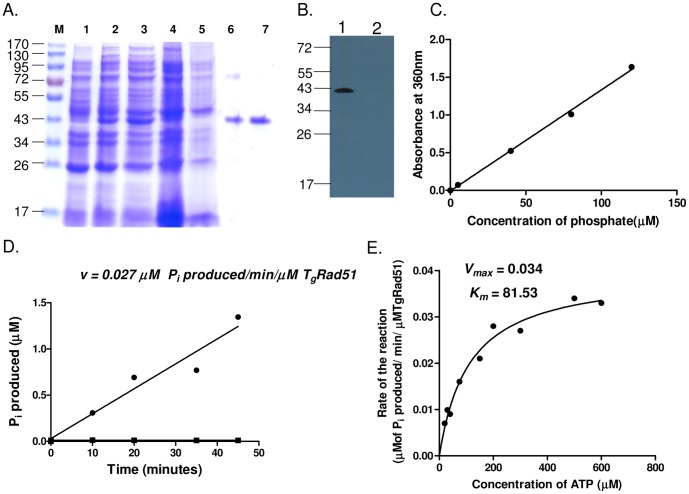
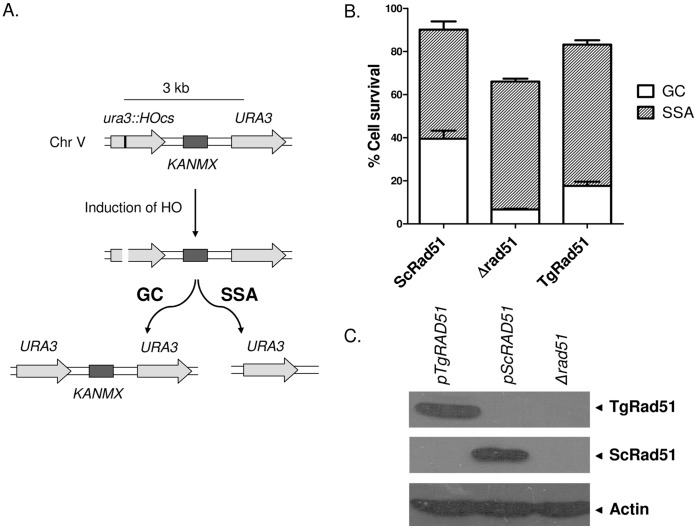
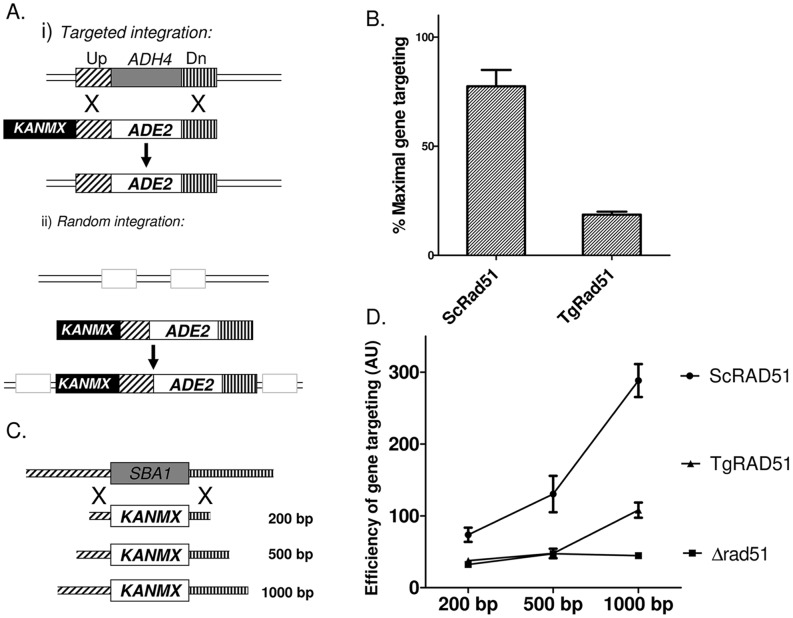
Repairing double strand breaks (DSBs) is absolutely essential for the survival of obligate intracellular parasite Toxoplasma gondii. Thus, DSB repair mechanisms could be excellent targets for chemotherapeutic interventions. Recent genetic and bioinformatics analyses confirm the presence of both homologous recombination (HR) as well as non homologous end joining (NHEJ) proteins in this lower eukaryote. In order to get mechanistic insights into the HR mediated DSB repair pathway in this parasite, we have characterized the key protein involved in homologous recombination, namely TgRad51, at the biochemical and genetic levels. We have purified recombinant TgRad51 protein to 99% homogeneity and have characterized it biochemically. The ATP hydrolysis activity of TgRad51 shows a higher K(M) and much lower k(cat) compared to bacterial RecA or Rad51 from other related protozoan parasites. Taking yeast as a surrogate model system we have shown that TgRad51 is less efficient in gene conversion mechanism. Further, we have found that TgRad51 mediated gene integration is more prone towards random genetic loci rather than targeted locus. We hypothesize that compromised ATPase activity of TgRad51 is responsible for inefficient gene targeting and poor gene conversion efficiency in this protozoan parasite. With increase in homologous flanking regions almost three fold increments in targeted gene integration is observed, which is similar to the trend found with ScRad51. Our findings not only help us in understanding the reason behind inefficient gene targeting in T. gondii but also could be exploited to facilitate high throughput knockout as well as epitope tagging of Toxoplasma genes.
修复双链断裂(DSB)对于专性细胞内寄生虫刚地弓形虫的生存是绝对必要的。因此,DSB 修复机制可能是化学治疗干预的极好靶点。最近的遗传和生物信息学分析证实,这种较低等的真核生物中存在同源重组(HR)和非同源末端连接(NHEJ)蛋白。为了深入了解该寄生虫中 HR 介导的 DSB 修复途径的机制,我们在生化和遗传水平上对参与同源重组的关键蛋白 TgRad51 进行了表征。我们已经将重组 TgRad51 蛋白纯化至 99%的纯度,并对其进行了生化表征。与细菌 RecA 或来自其他相关原生动物寄生虫的 Rad51 相比,TgRad51 的 ATP 水解活性显示出更高的 K(M)和低得多的 k(cat)。我们以酵母为替代模型系统,表明 TgRad51 在基因转换机制中的效率较低。此外,我们发现 TgRad51 介导的基因整合更容易发生在随机遗传基因座,而不是靶向基因座。我们假设 TgRad51 的 ATP 酶活性受损是导致该原生动物寄生虫基因靶向效率低下和基因转换效率差的原因。随着同源侧翼区域的增加,靶向基因整合增加了近三倍,这与 ScRad51 发现的趋势相似。我们的研究结果不仅帮助我们了解了 T. gondii 中基因靶向效率低下的原因,还可以利用这些结果来促进高效的基因敲除以及弓形虫基因的表位标记。