Dalton Cardiovascular Research Center, University of Missouri-Columbia, Columbia, MO 65211, USA.
J Gen Physiol. 2012 Oct;140(4):347-59. doi: 10.1085/jgp.201210834. Epub 2012 Sep 10.
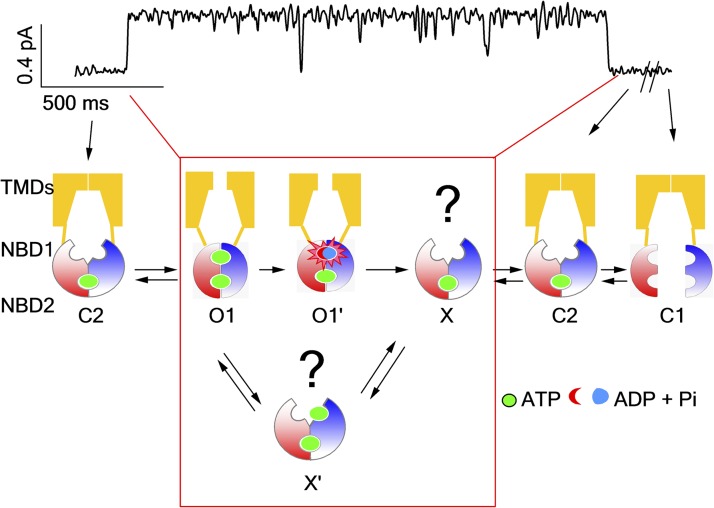
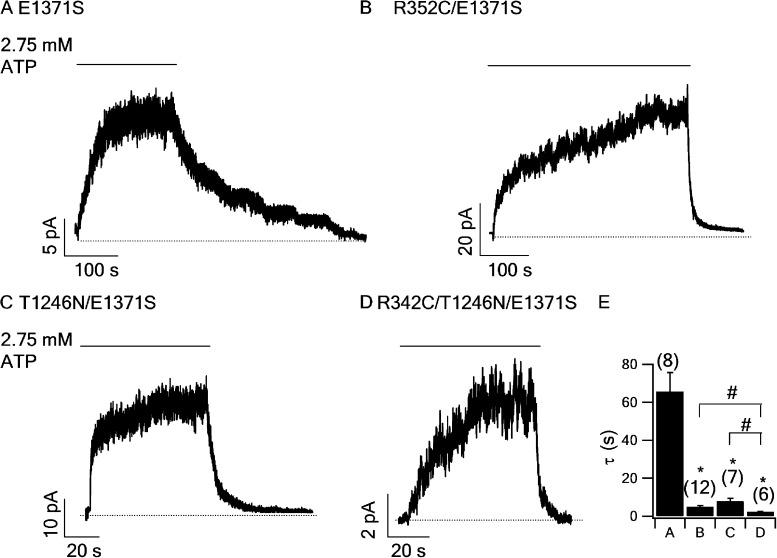
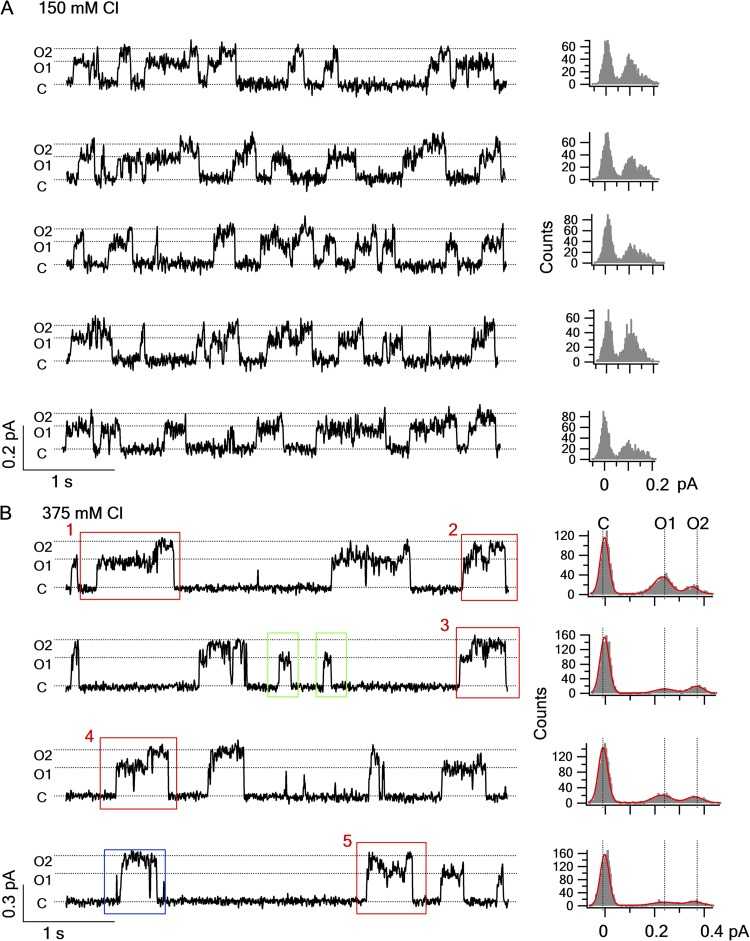
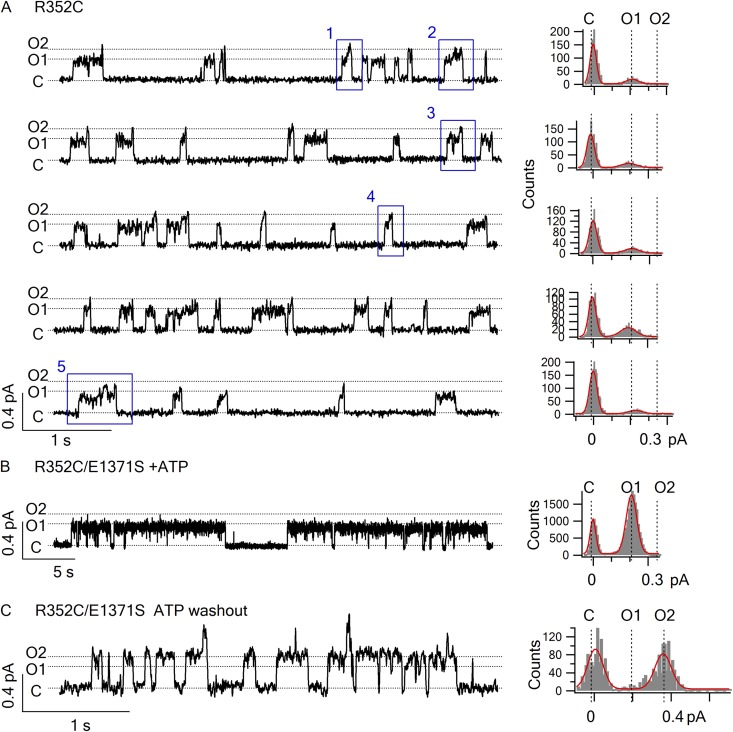
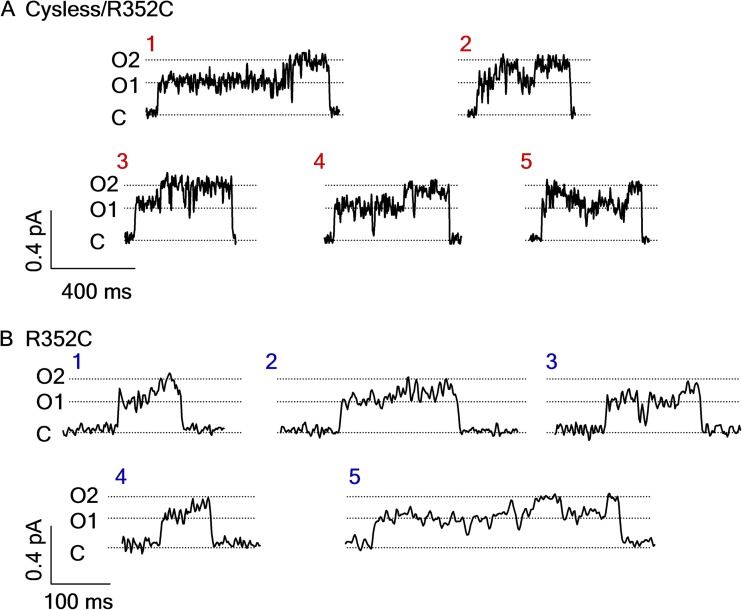
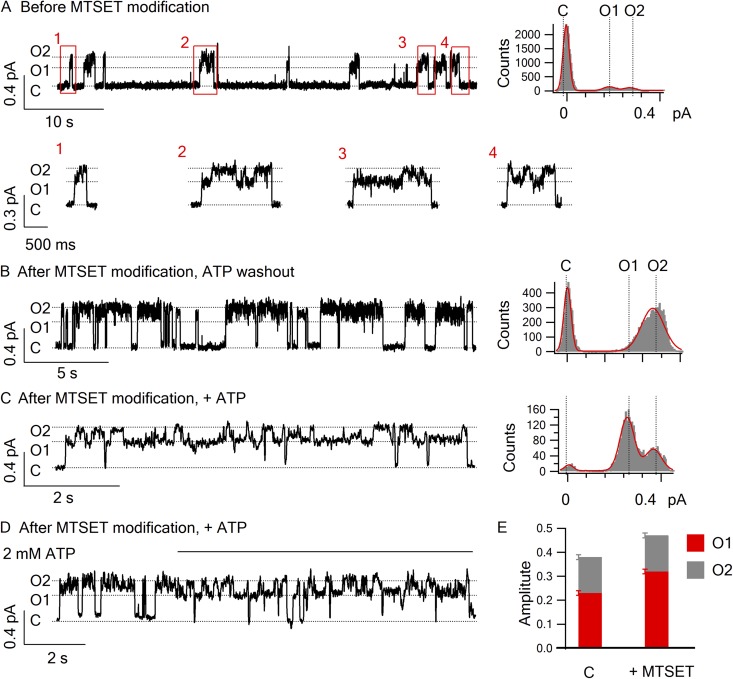
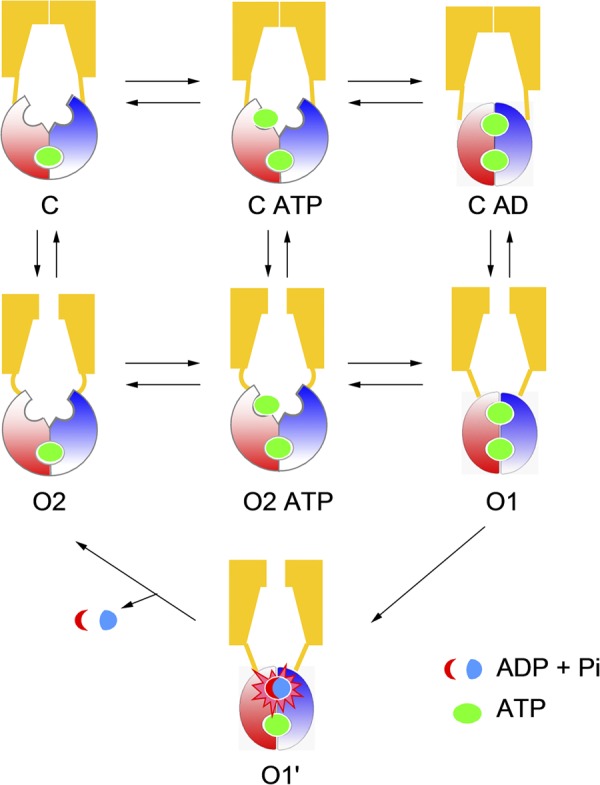
Cystic fibrosis transmembrane conductance regulator (CFTR) is a unique member of the ATP-binding cassette (ABC) protein superfamily. Unlike most other ABC proteins that function as active transporters, CFTR is an ATP-gated chloride channel. The opening of CFTR's gate is associated with ATP-induced dimerization of its two nucleotide-binding domains (NBD1 and NBD2), whereas gate closure is facilitated by ATP hydrolysis-triggered partial separation of the NBDs. This generally held theme of CFTR gating-a strict coupling between the ATP hydrolysis cycle and the gating cycle-is put to the test by our recent finding of a short-lived, post-hydrolytic state that can bind ATP and reenter the ATP-induced original open state. We accidentally found a mutant CFTR channel that exhibits two distinct open conductance states, the smaller O1 state and the larger O2 state. In the presence of ATP, the transition between the two states follows a preferred O1→O2 order, a telltale sign of a violation of microscopic reversibility, hence demanding an external energy input likely from ATP hydrolysis, as such preferred gating transition was abolished in a hydrolysis-deficient mutant. Interestingly, we also observed a considerable amount of opening events that contain more than one O1→O2 transition, indicating that more than one ATP molecule may be hydrolyzed within an opening burst. We thus conclude a nonintegral stoichiometry between the gating cycle and ATP consumption. Our results lead to a six-state gating model conforming to the classical allosteric mechanism: both NBDs and transmembrane domains hold a certain degree of autonomy, whereas the conformational change in one domain will facilitate the conformational change in the other domain.
囊性纤维化跨膜电导调节因子(CFTR)是 ATP 结合盒(ABC)蛋白超家族的独特成员。与大多数其他作为主动转运体发挥作用的 ABC 蛋白不同,CFTR 是一种 ATP 门控氯离子通道。CFTR 门的打开与两个核苷酸结合域(NBD1 和 NBD2)的 ATP 诱导二聚化有关,而门的关闭则由 ATP 水解触发的 NBD 部分分离来促进。CFTR 门控的这一普遍主题——ATP 水解循环与门控循环之间的严格偶联——受到我们最近发现的短暂、水解后状态的考验,该状态可以结合 ATP 并重新进入 ATP 诱导的原始开放状态。我们偶然发现了一种突变 CFTR 通道,它表现出两种不同的开放电导状态,较小的 O1 状态和较大的 O2 状态。在 ATP 存在的情况下,两种状态之间的转变遵循优先 O1→O2 顺序,这是违反微观可逆性的明显迹象,因此需要外部能量输入,可能来自 ATP 水解,因为这种优先门控转变在水解缺陷突变体中被消除。有趣的是,我们还观察到相当数量的开放事件包含不止一次 O1→O2 转变,表明在开放爆发中可能水解不止一个 ATP 分子。因此,我们得出结论,门控循环和 ATP 消耗之间的非整数计量关系。我们的结果导致了一个符合经典变构机制的六态门控模型:NBD 和跨膜结构域都具有一定程度的自主性,而一个结构域的构象变化将促进另一个结构域的构象变化。