Dept. of Bioscience and Informatics, Keio University, Yokohama, Japan.
PLoS One. 2012;7(12):e51000. doi: 10.1371/journal.pone.0051000. Epub 2012 Dec 20.
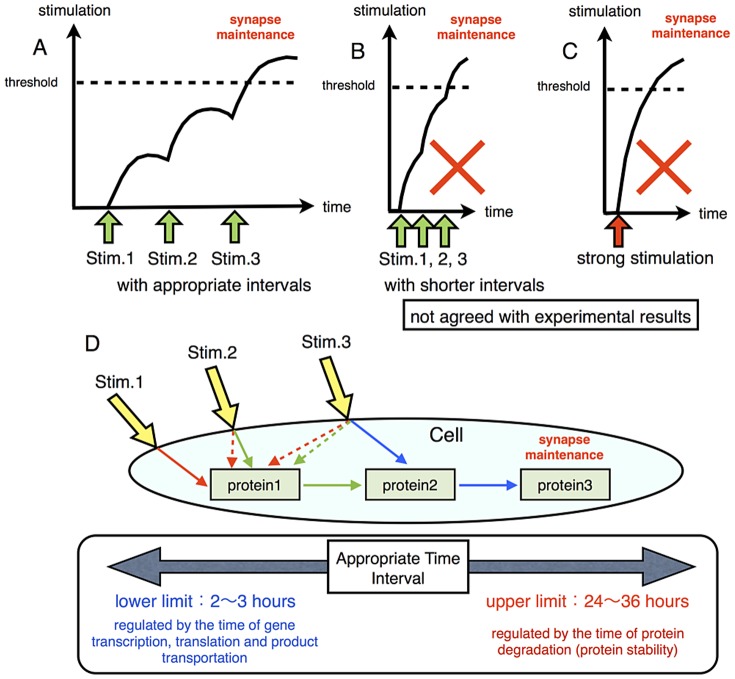
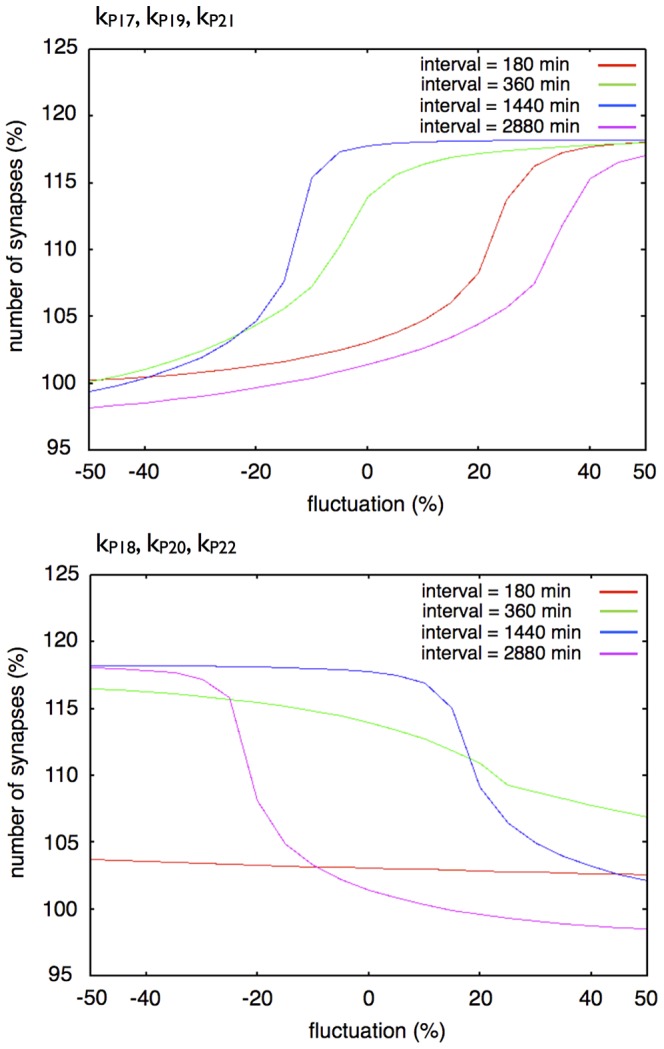
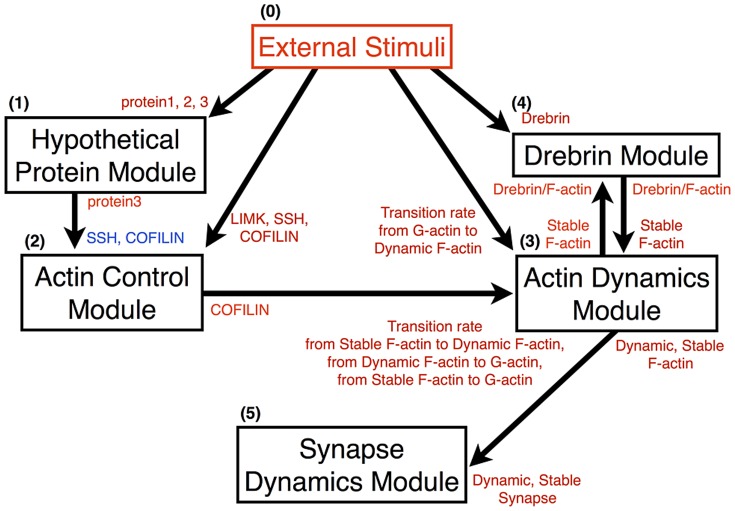
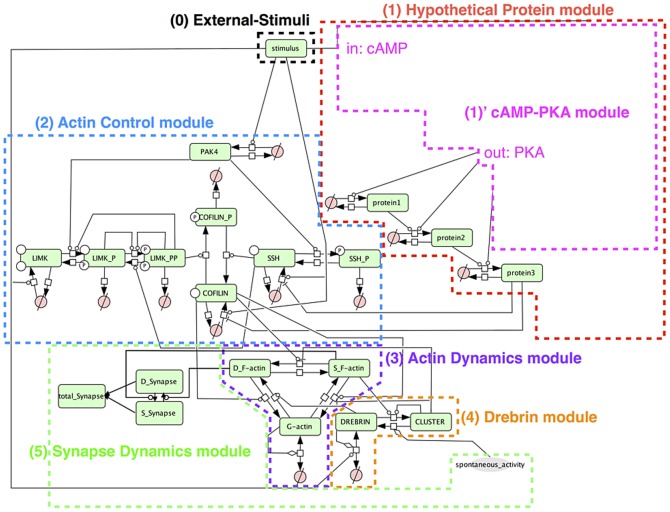
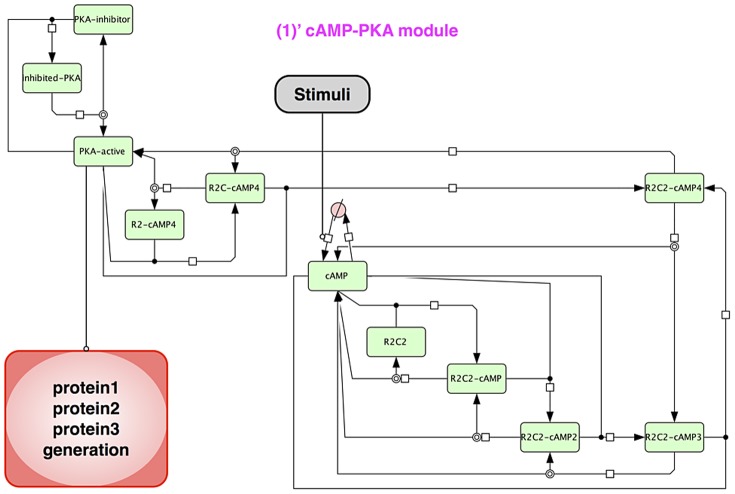
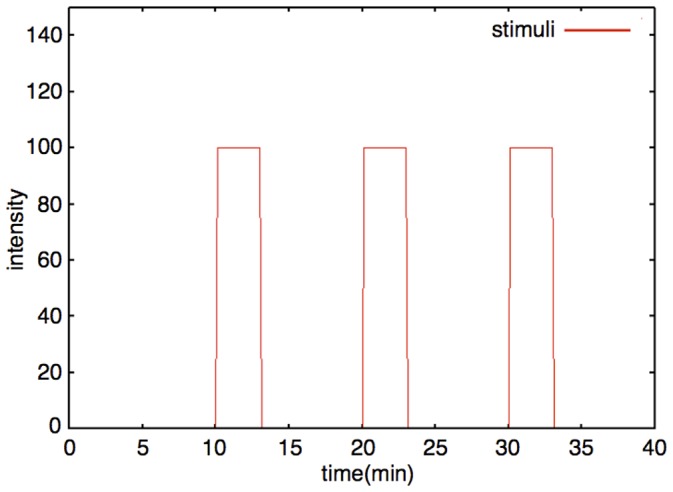
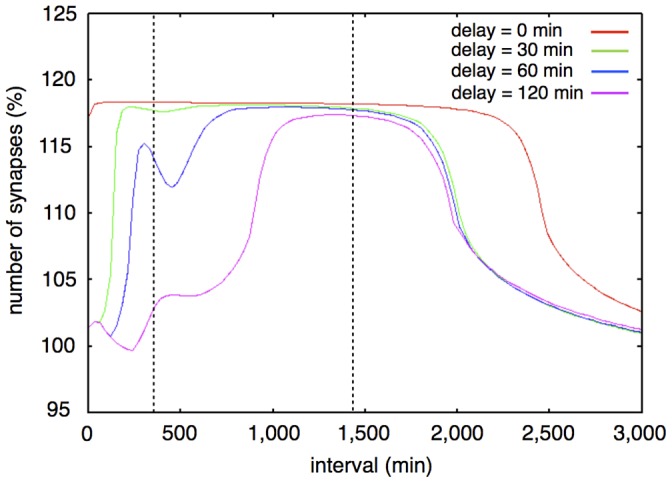
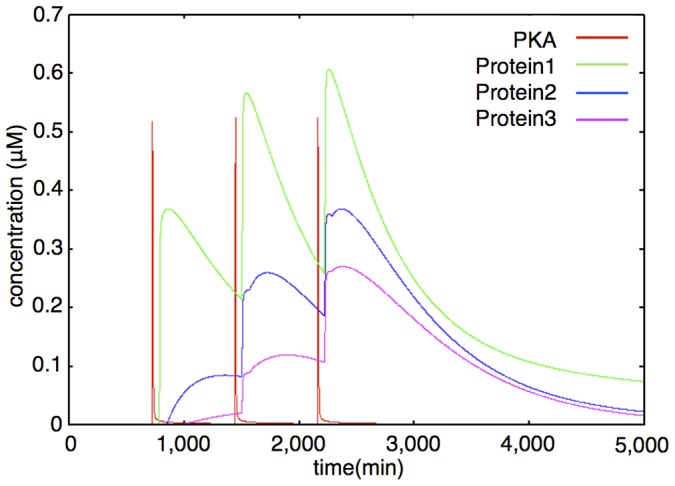
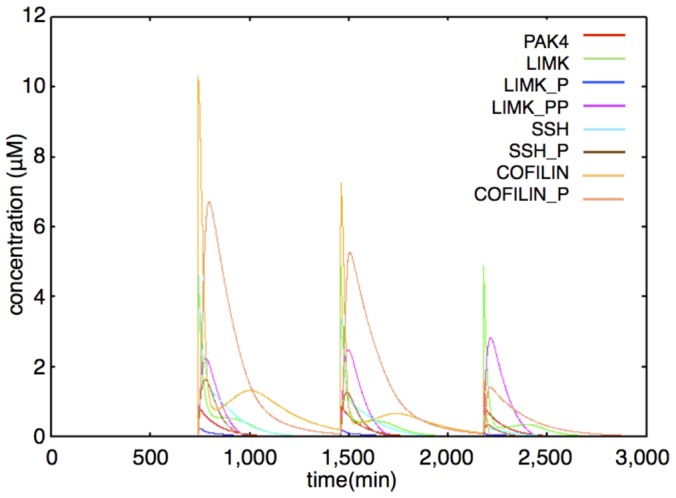
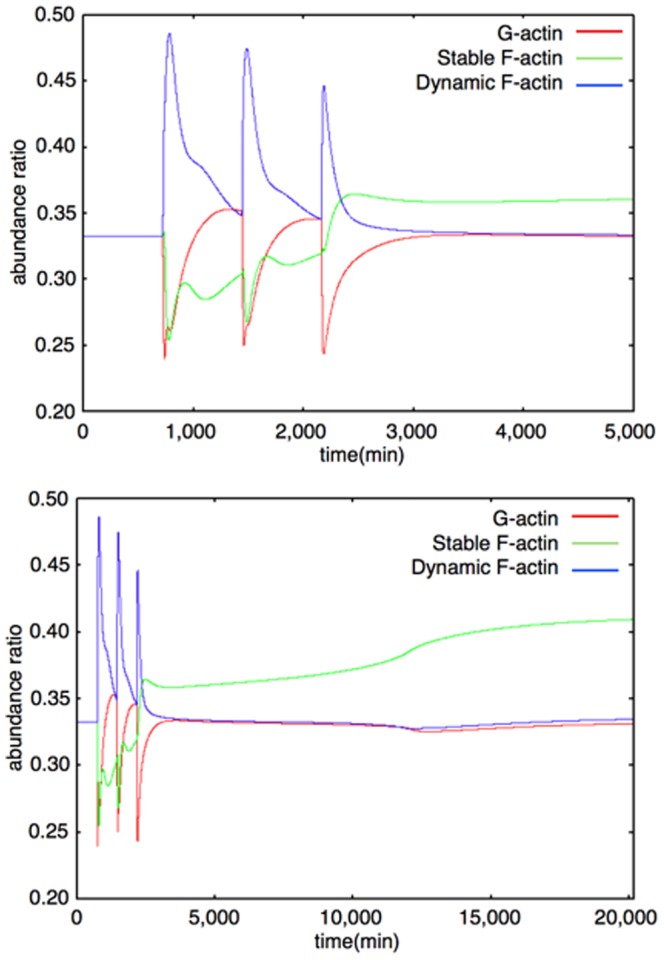
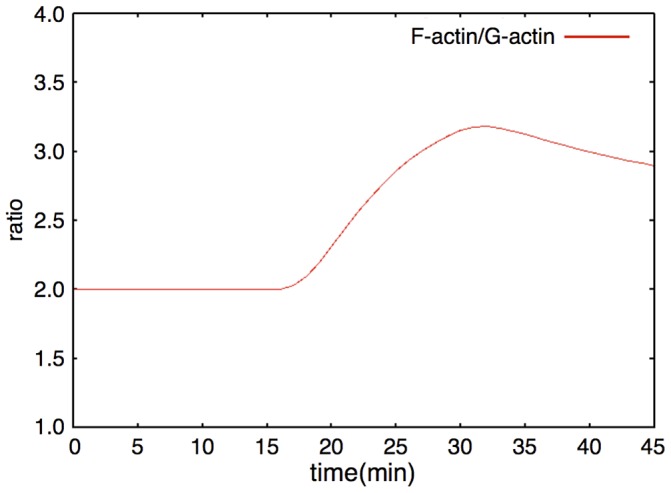
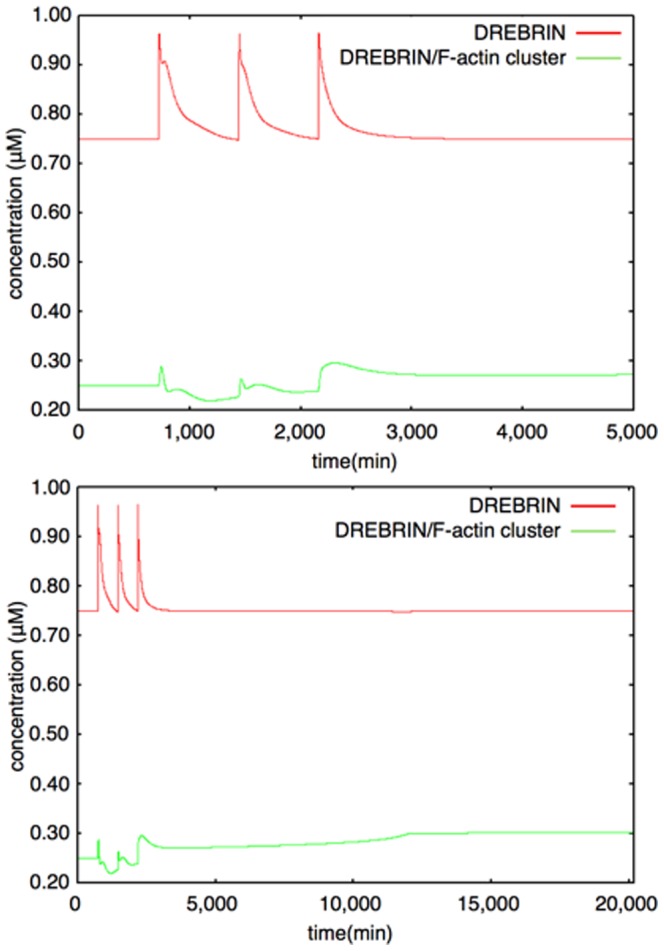
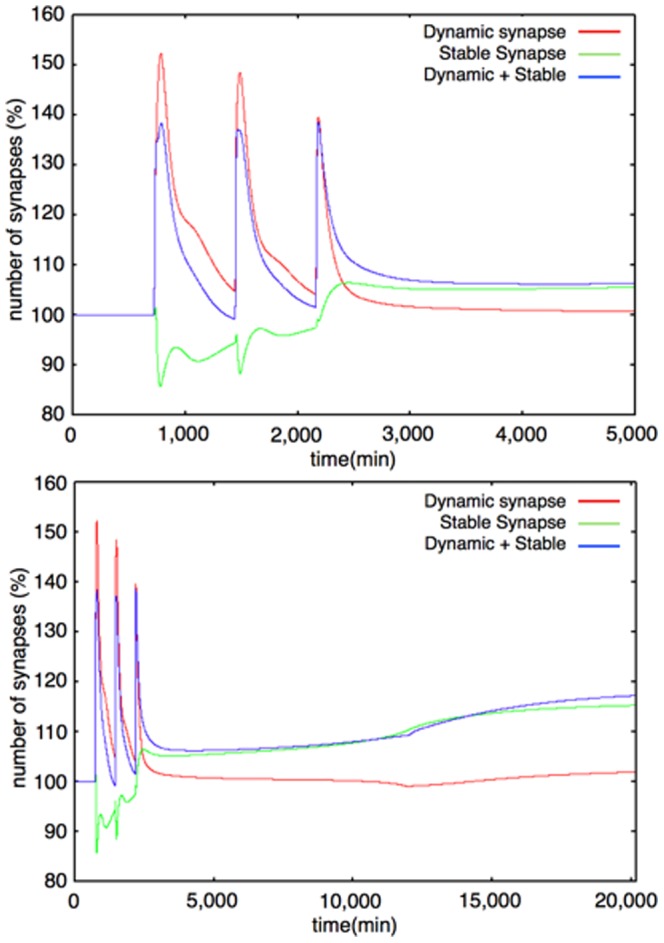
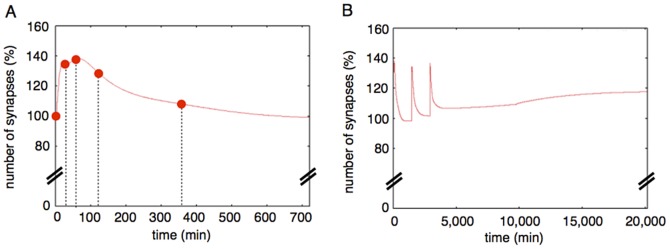
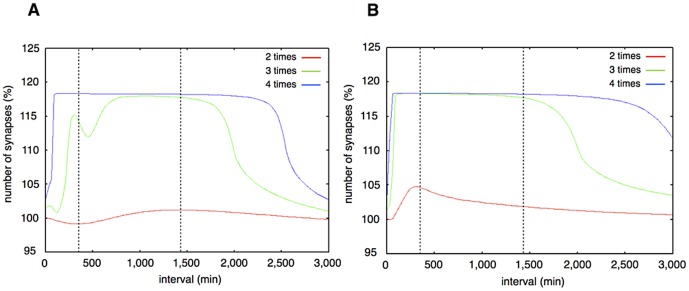
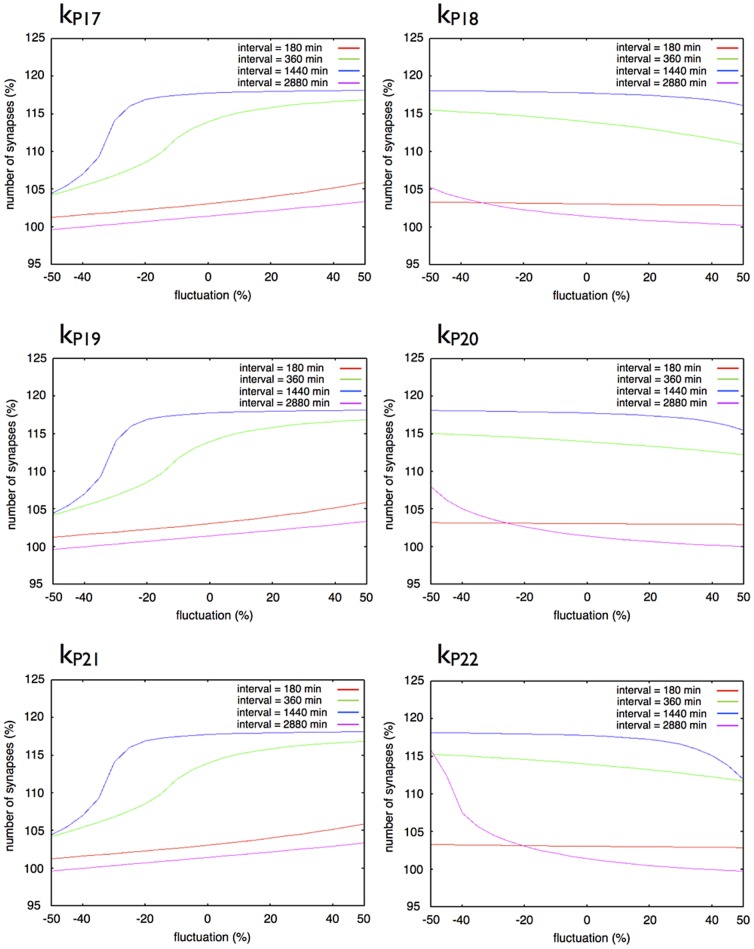
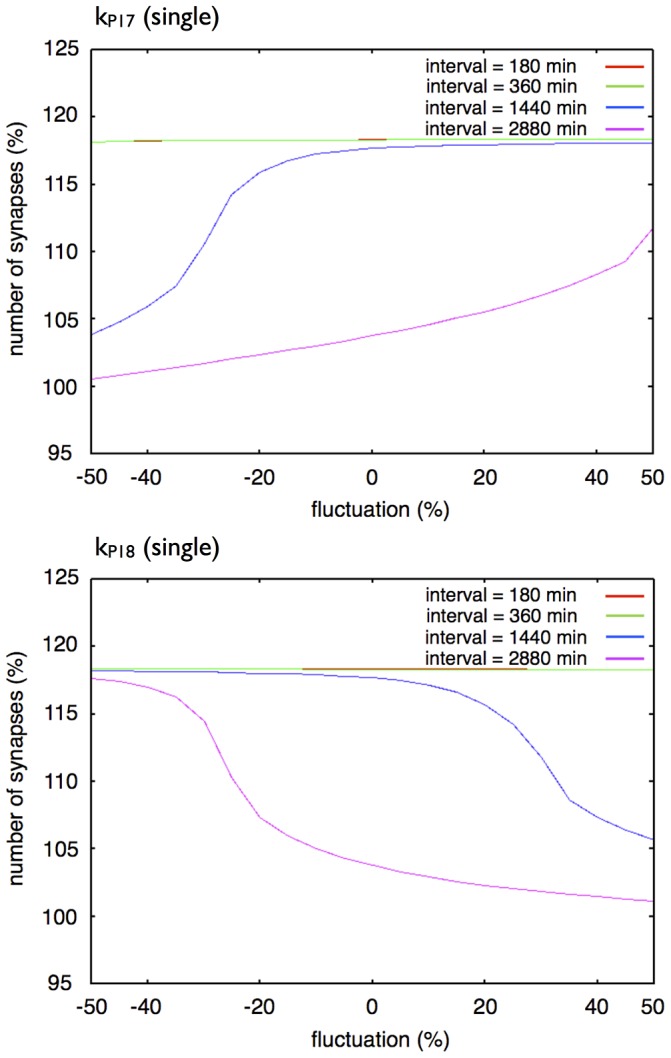
The mechanisms of long-term synaptic maintenance are a key component to understanding the mechanism of long-term memory. From biological experiments, a hypothesis arose that repetitive stimuli with appropriate intervals are essential to maintain new synapses for periods of longer than a few days. We successfully reproduce the time-course of relative numbers of synapses with our mathematical model in the same conditions as biological experiments, which used Adenosine-3', 5'-cyclic monophosphorothioate, Sp-isomer (Sp-cAMPS) as external stimuli. We also reproduce synaptic maintenance responsiveness to intervals of Sp-cAMPS treatment accompanied by PKA activation. The model suggests a possible mechanism of sustainable synaptogenesis which consists of two steps. First, the signal transduction from an external stimulus triggers the synthesis of a new signaling protein. Second, the new signaling protein is required for the next signal transduction with the same stimuli. As a result, the network component is modified from the first network, and a different signal is transferred which triggers the synthesis of another new signaling molecule. We refer to this hypothetical mechanism as network succession. We build our model on the basis of two hypotheses: (1) a multi-step network succession induces downregulation of SSH and COFILIN gene expression, which triggers the production of stable F-actin; (2) the formation of a complex of stable F-actin with Drebrin at PSD is the critical mechanism to achieve long-term synaptic maintenance. Our simulation shows that a three-step network succession is sufficient to reproduce sustainable synapses for a period longer than 14 days. When we change the network structure to a single step network, the model fails to follow the exact condition of repetitive signals to reproduce a sufficient number of synapses. Another advantage of the three-step network succession is that this system indicates a greater tolerance of parameter changes than the single step network.
长期突触维持的机制是理解长期记忆机制的关键组成部分。从生物学实验中,出现了一个假设,即具有适当间隔的重复刺激对于维持新突触的时间超过几天是必不可少的。我们使用腺嘌呤 3',5'-环单磷酸硫代酯,Sp-异构体(Sp-cAMPS)作为外部刺激,在相同的条件下成功地用我们的数学模型再现了相对突触数量的时间过程。我们还再现了突触维持对 Sp-cAMPS 处理间隔的反应性,同时伴有 PKA 激活。该模型提出了一种可持续突触发生的可能机制,该机制由两个步骤组成。首先,外部刺激的信号转导触发新信号蛋白的合成。其次,新信号蛋白是下一个具有相同刺激的信号转导所必需的。因此,网络组件从第一个网络中修改,并且传递不同的信号,触发另一种新信号分子的合成。我们将这种假设的机制称为网络连续。我们基于两个假设构建我们的模型:(1)多步网络连续诱导 SSH 和 COFILIN 基因表达的下调,从而触发稳定的 F-肌动蛋白的产生;(2)稳定的 F-肌动蛋白与 PSD 处的 Drebrin 形成复合物是实现长期突触维持的关键机制。我们的模拟表明,三步网络连续足以再现持续超过 14 天的突触。当我们将网络结构更改为单步网络时,模型无法根据重复信号的精确条件再现足够数量的突触。三步网络连续的另一个优点是,该系统比单步网络具有更大的参数变化容忍度。