Laboratory of Chemical Physics, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892, USA.
Acc Chem Res. 2013 Jul 16;46(7):1487-96. doi: 10.1021/ar300282r. Epub 2013 Jan 7.
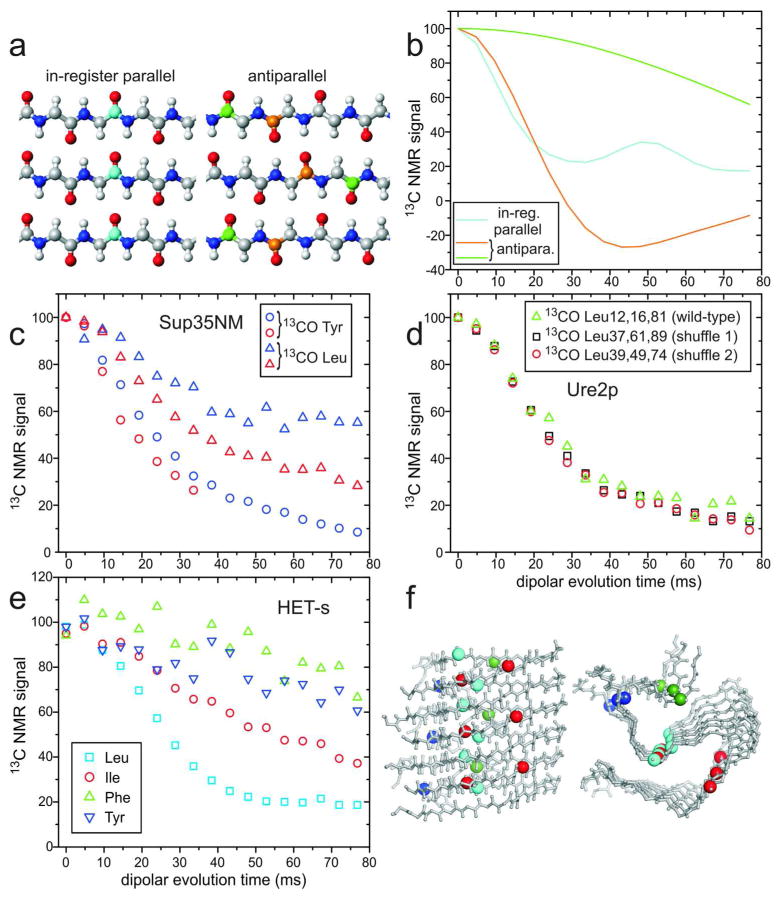
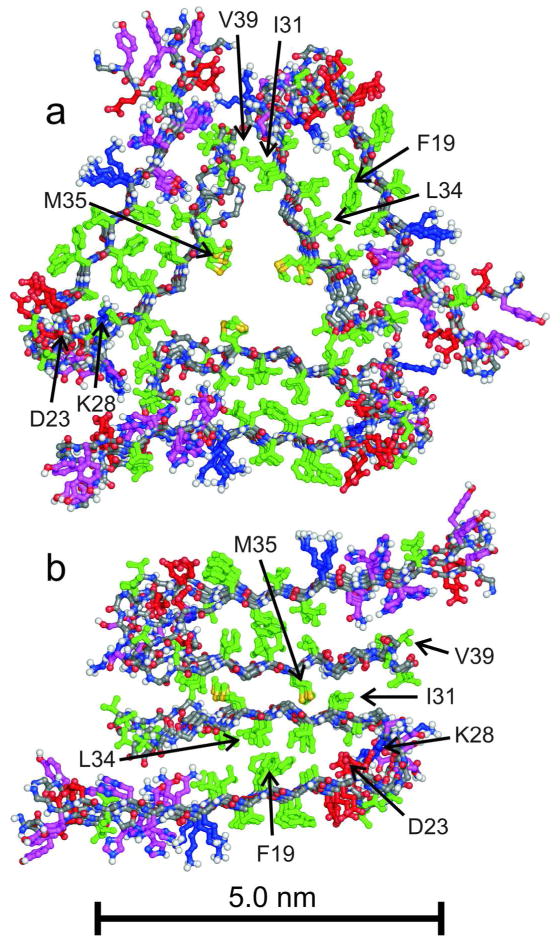
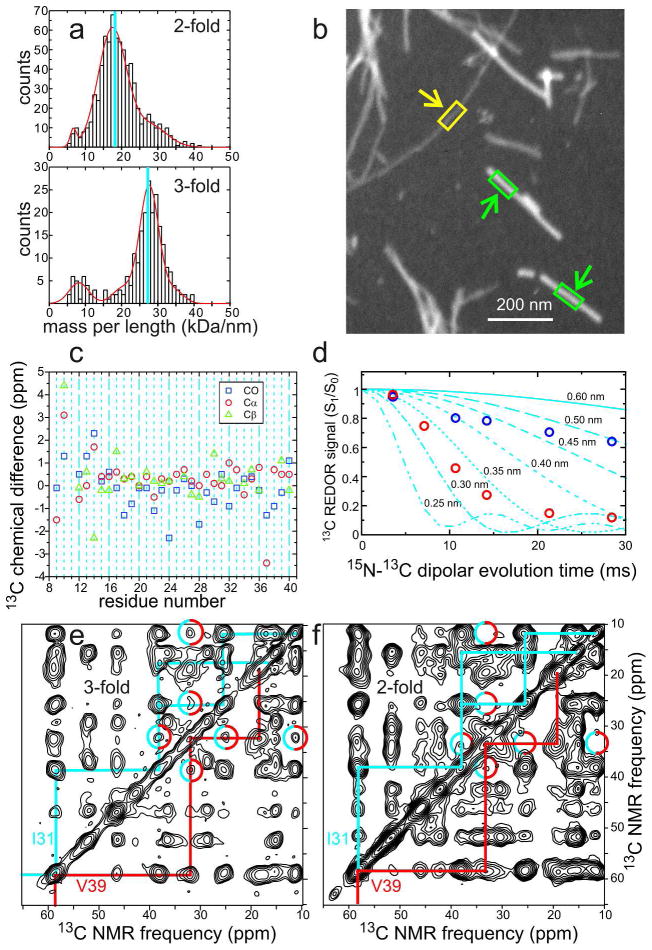
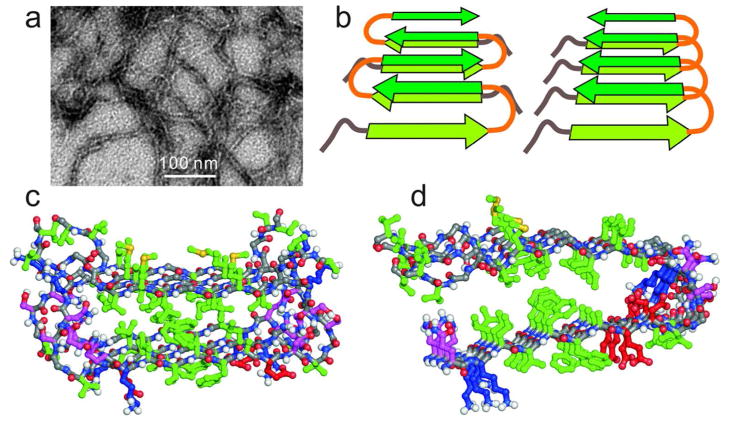
Many peptides and proteins self-assemble into amyloid fibrils. Examples include mammalian and fungal prion proteins, polypeptides associated with human amyloid diseases, and proteins that may have biologically functional amyloid states. To understand the propensity for polypeptides to form amyloid fibrils and to facilitate rational design of amyloid inhibitors and imaging agents, it is necessary to elucidate the molecular structures of these fibrils. Although fibril structures were largely mysterious 15 years ago, a considerable body of reliable structural information about amyloid fibril structures now exists, with essential contributions from solid state nuclear magnetic resonance (NMR) measurements. This Account reviews results from our laboratories and discusses several structural issues that have been controversial. In many cases, the amino acid sequences of amyloid fibrils do not uniquely determine their molecular structures. Self-propagating, molecular-level polymorphism complicates the structure determination problem and can lead to apparent disagreements between results from different laboratories, particularly when different laboratories study different polymorphs. For 40-residue β-amyloid (Aβ₁₋₄₀) fibrils associated with Alzheimer's disease, we have developed detailed structural models from solid state NMR and electron microscopy data for two polymorphs. These polymorphs have similar peptide conformations, identical in-register parallel β-sheet organizations, but different overall symmetry. Other polymorphs have also been partially characterized by solid state NMR and appear to have similar structures. In contrast, cryo-electron microscopy studies that use significantly different fibril growth conditions have identified structures that appear (at low resolution) to be different from those examined by solid state NMR. Based on solid state NMR and electron paramagnetic resonance (EPR) measurements, the in-register parallel β-sheet organization found in β-amyloid fibrils also occurs in many other fibril-forming systems. We attribute this common structural motif to the stabilization of amyloid structures by intermolecular interactions among like amino acids, including hydrophobic interactions and polar zippers. Surprisingly, we have recently identified and characterized antiparallel β-sheets in certain fibrils that are formed by the D23N mutant of Aβ₁₋₄₀, a mutant that is associated with early-onset, familial neurodegenerative disease. Antiparallel D23N-Aβ₁₋₄₀ fibrils are metastable with respect to parallel structures and, therefore, represent an off-pathway intermediate in the amyloid fibril formation process. Other methods have recently produced additional evidence for antiparallel β-sheets in other amyloid-formation intermediates. As an alternative to simple parallel and antiparallel β-sheet structures, researchers have proposed β-helical structural models for some fibrils, especially those formed by mammalian and fungal prion proteins. Solid state NMR and EPR data show that fibrils formed in vitro by recombinant PrP have in-register parallel β-sheet structures. However, the structure of infectious PrP aggregates is not yet known. The fungal HET-s prion protein has been shown to contain a β-helical structure. However, all yeast prions studied by solid state NMR (Sup35p, Ure2p, and Rnq1p) have in-register parallel β-sheet structures, with their Gln- and Asn-rich N-terminal segments forming the fibril core.
许多肽和蛋白质会自组装成淀粉样纤维。哺乳动物和真菌朊病毒蛋白、与人类淀粉样疾病相关的多肽,以及可能具有生物学功能的淀粉样状态的蛋白质都属于此类。为了了解多肽形成淀粉样纤维的倾向,并促进淀粉样抑制剂和成像剂的合理设计,有必要阐明这些纤维的分子结构。尽管 15 年前纤维的结构在很大程度上还是个谜,但现在已经存在大量关于淀粉样纤维结构的可靠结构信息,其中固态核磁共振(NMR)测量做出了重要贡献。本综述回顾了我们实验室的研究结果,并讨论了几个存在争议的结构问题。在许多情况下,淀粉样纤维的氨基酸序列并不能唯一确定其分子结构。自传播的分子级别的多态性使结构确定问题变得复杂,并可能导致不同实验室之间的结果出现明显分歧,特别是当不同实验室研究不同的多态体时。对于与阿尔茨海默病相关的 40 个残基β-淀粉样蛋白(Aβ₁₋₄₀)纤维,我们根据固态 NMR 和电子显微镜数据为两种多态体开发了详细的结构模型。这些多态体具有相似的肽构象,相同的规则平行β-折叠组织,但整体对称性不同。其他多态体也已通过固态 NMR 进行了部分表征,似乎具有相似的结构。相比之下,使用差异显著的纤维生长条件的低温电子显微镜研究鉴定出的结构(在低分辨率下)似乎与固态 NMR 检查的结构不同。基于固态 NMR 和电子顺磁共振(EPR)测量,β-淀粉样纤维中发现的规则平行β-折叠组织也存在于许多其他纤维形成系统中。我们将这种共同的结构基序归因于相似氨基酸之间的分子间相互作用对淀粉样结构的稳定作用,包括疏水相互作用和极性拉链。令人惊讶的是,我们最近在由 Aβ₁₋₄₀的 D23N 突变体形成的某些纤维中鉴定并表征了反平行β-折叠,该突变体与早发性家族性神经退行性疾病有关。反平行 D23N-Aβ₁₋₄₀纤维相对于平行结构是亚稳定的,因此代表淀粉样纤维形成过程中的一种非途径中间体。其他方法最近也为其他淀粉样形成中间体中的反平行β-折叠提供了额外的证据。作为简单的平行和反平行β-折叠结构的替代方案,研究人员提出了一些纤维的β-螺旋结构模型,特别是那些由哺乳动物和真菌朊病毒蛋白形成的纤维。固态 NMR 和 EPR 数据表明,在体外由重组 PrP 形成的纤维具有规则平行的β-折叠结构。然而,传染性 PrP 聚集体的结构仍未知。真菌 HET-s 朊病毒蛋白已被证明含有β-螺旋结构。然而,通过固态 NMR 研究的所有酵母朊病毒(Sup35p、Ure2p 和 Rnq1p)都具有规则平行的β-折叠结构,其富含 Gln 和 Asn 的 N 端片段构成纤维核心。