Laboratory of Microbiology, Department of Agrotechnology and Food Sciences, Wageningen University, Wageningen, The Netherlands.
RNA Biol. 2013 May;10(5):749-61. doi: 10.4161/rna.24202. Epub 2013 Mar 27.
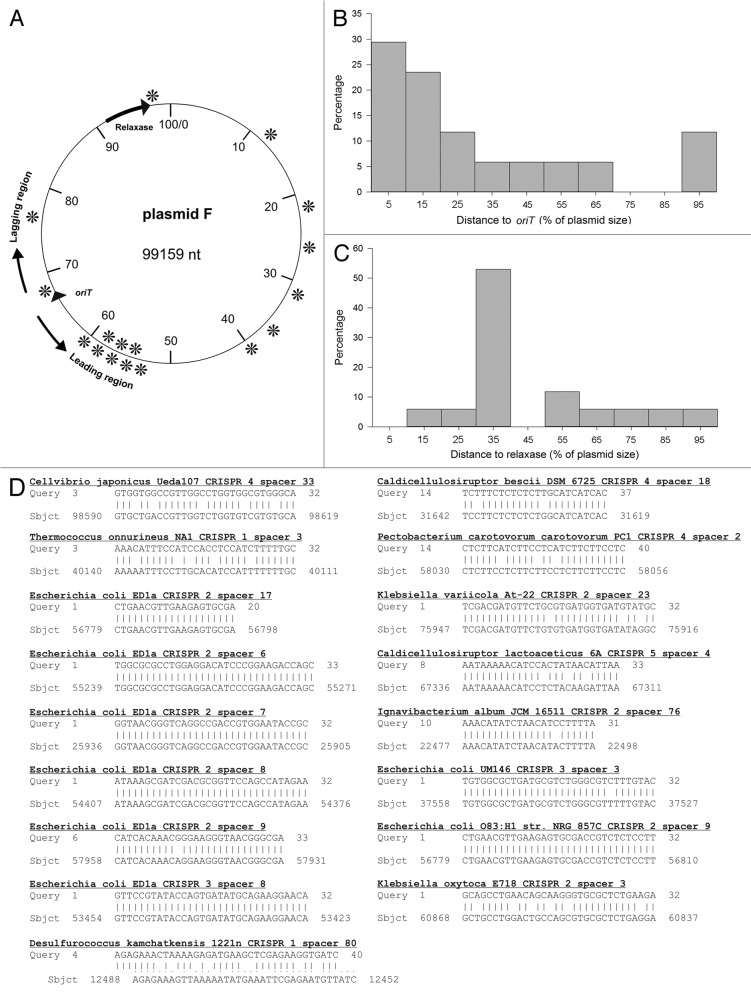
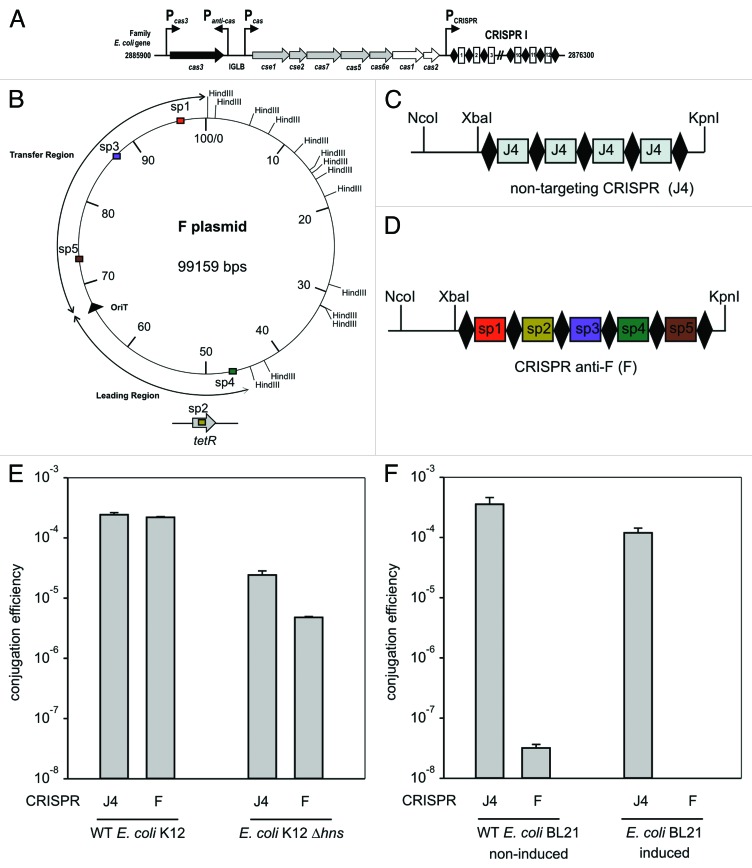
Most prokaryotes contain CRISPR-Cas immune systems that provide protection against mobile genetic elements. We have focused on the ability of CRISPR-Cas to block plasmid conjugation, and analyzed the position of target sequences (protospacers) on conjugative plasmids. The analysis reveals that protospacers are non-uniformly distributed over plasmid regions in a pattern that is determined by the plasmid's mobilization type (MOB). While MOBP plasmids are most frequently targeted in the region entering the recipient cell last (lagging region), MOBF plasmids are mostly targeted in the region entering the recipient cell first (leading region). To explain this protospacer distribution bias, we propose two mutually non-exclusive hypotheses: (1) spacers are acquired more frequently from either the leading or lagging region depending on the MOB type (2) CRISPR-interference is more efficient when spacers target these preferred regions. To test the latter hypothesis, we analyzed Type I-E CRISPR-interference against MOBF prototype plasmid F in Escherichia coli. Our results show that plasmid conjugation is effectively inhibited, but the level of immunity is not affected by targeting the plasmid in the leading or lagging region. Moreover, CRISPR-immunity levels do not depend on whether the incoming single-stranded plasmid DNA, or the DNA strand synthesized in the recipient is targeted. Our findings indicate that single-stranded DNA may not be a target for Type I-E CRISPR-Cas systems, and suggest that the protospacer distribution bias might be due to spacer acquisition preferences.
大多数原核生物都含有 CRISPR-Cas 免疫系统,可提供针对移动遗传元件的保护。我们专注于 CRISPR-Cas 阻断质粒接合的能力,并分析了目标序列(原间隔区)在可接合质粒上的位置。分析表明,原间隔区在质粒区域中的分布不均匀,呈由质粒的移动类型(MOB)决定的模式。虽然 MOBP 质粒最常靶向进入受体细胞的最后一个区域(滞后区),但 MOBF 质粒主要靶向进入受体细胞的第一个区域(先导区)。为了解释这种原间隔区分布的偏向性,我们提出了两个相互排斥的假说:(1)根据 MOB 类型,间隔区从先导区或滞后区获得的频率更高;(2)当间隔区靶向这些首选区域时,CRISPR 干扰的效率更高。为了检验后一种假说,我们分析了 I-E 型 CRISPR 对大肠杆菌中 MOBF 原型质粒 F 的干扰。结果表明,质粒接合被有效抑制,但靶向先导区或滞后区对免疫水平没有影响。此外,CRISPR 免疫水平不受靶向进入的单链质粒 DNA 或受体中合成的 DNA 链的影响。我们的发现表明,单链 DNA 可能不是 I-E 型 CRISPR-Cas 系统的靶标,并提示原间隔区分布偏向可能是由于间隔区获得的偏好。