Zhao Yingying, Macgurn Jason A, Liu Max, Emr Scott
Weill Institute of Cell and Molecular Biology , Cornell University , Ithaca , United States.
Elife. 2013 Apr 16;2:e00459. doi: 10.7554/eLife.00459.
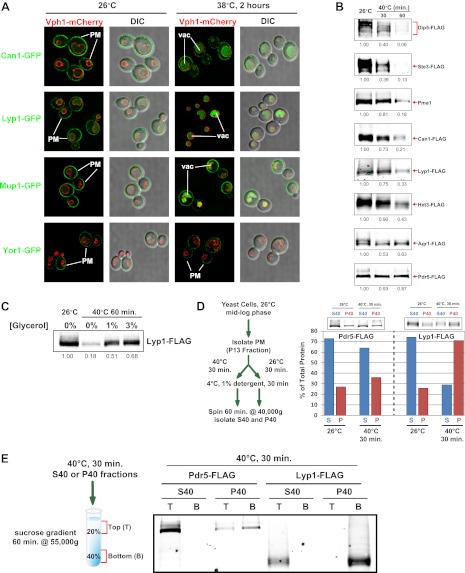
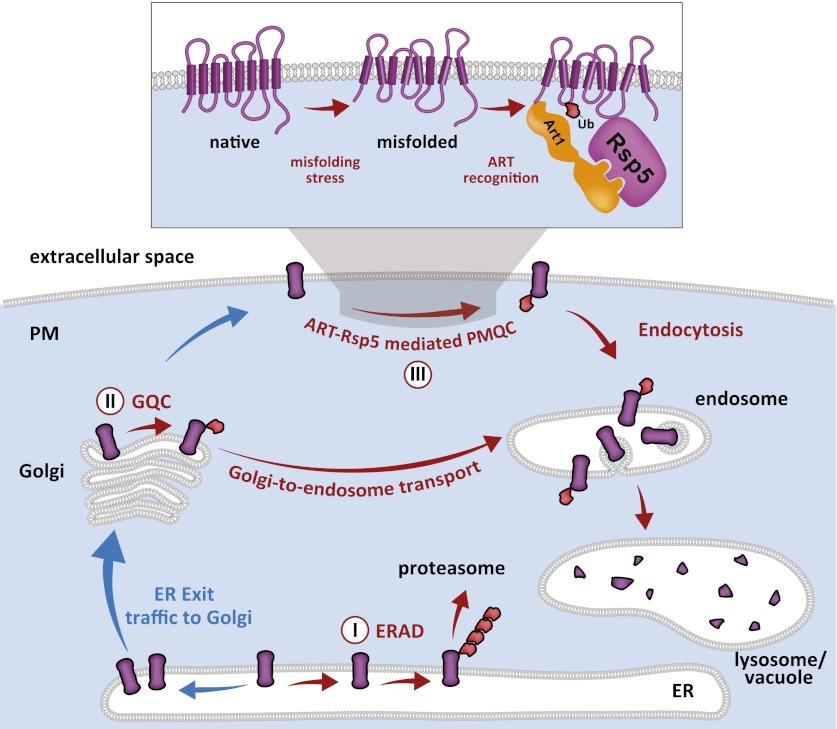
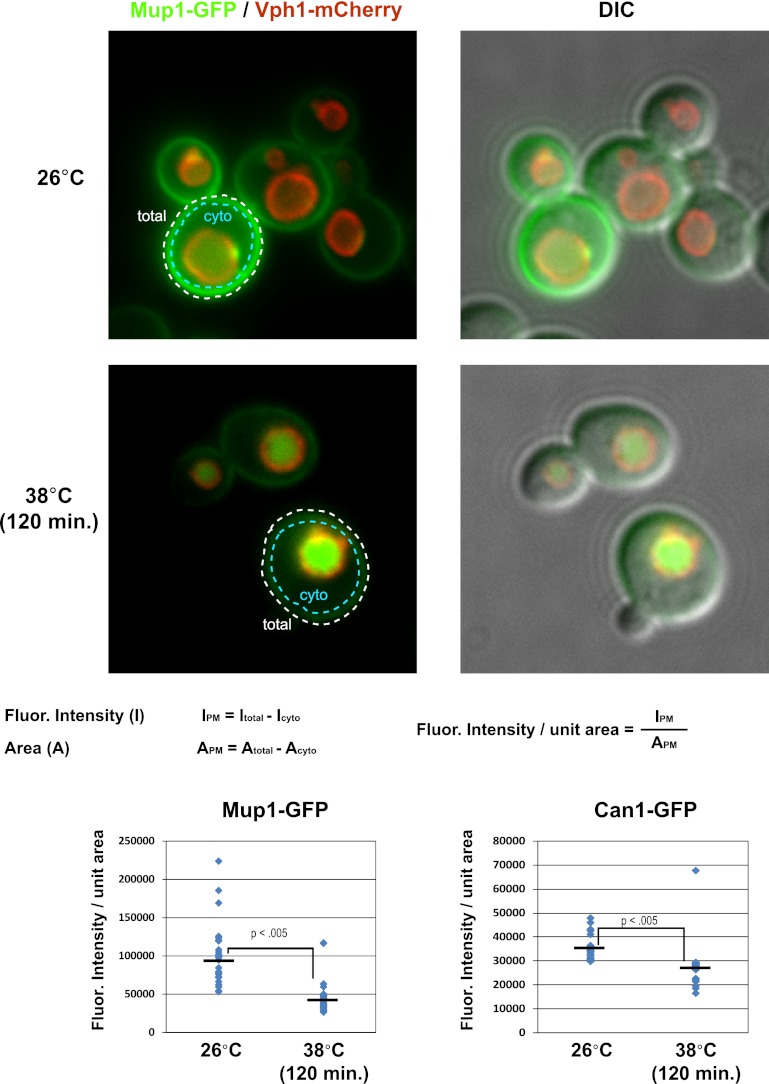
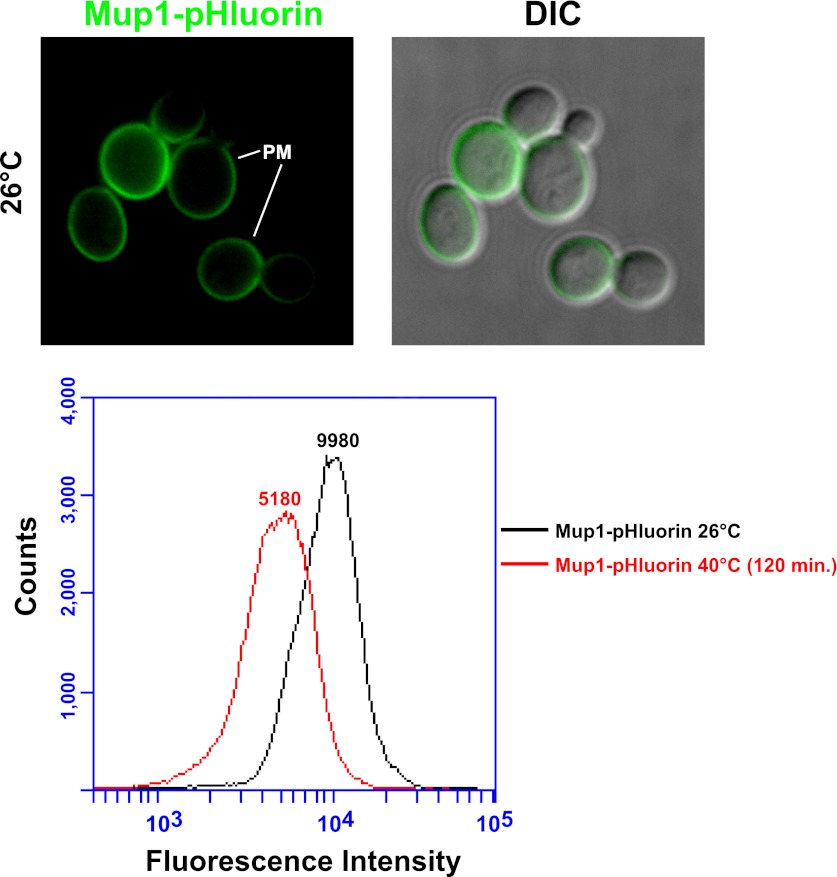
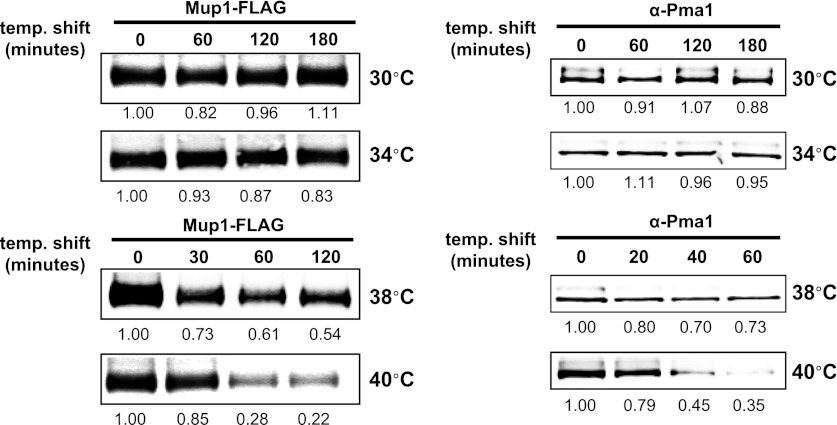
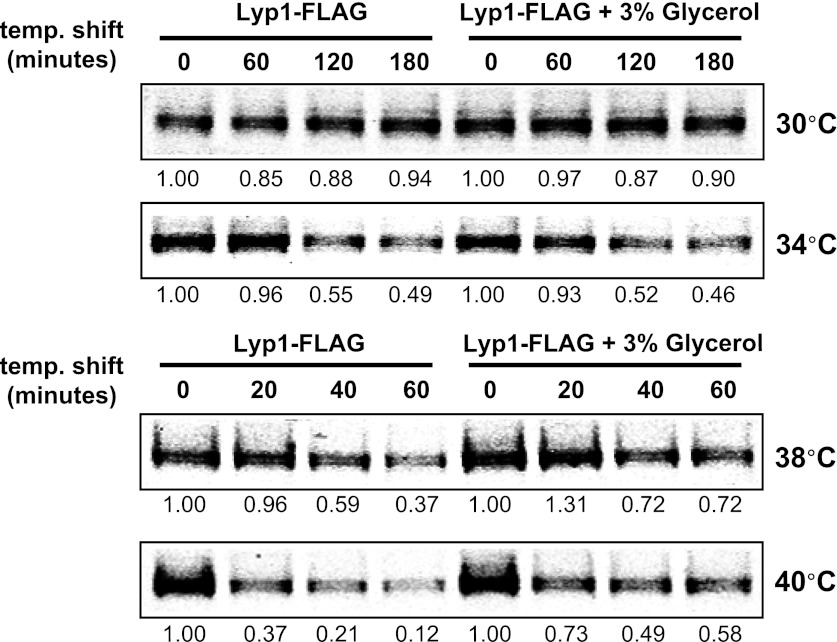
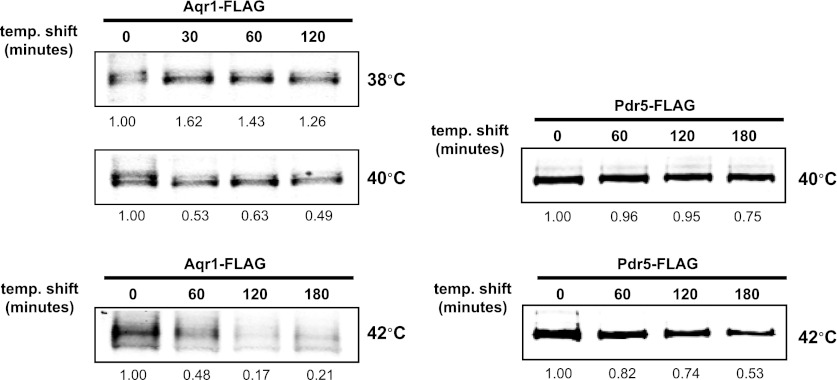
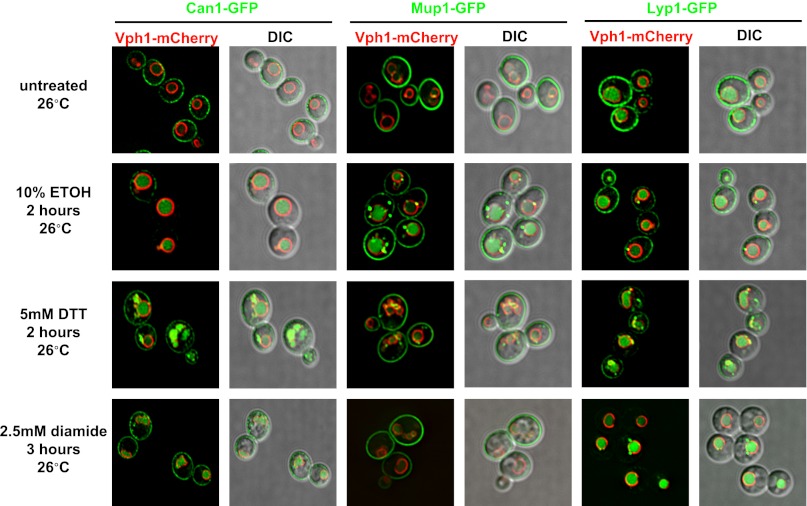
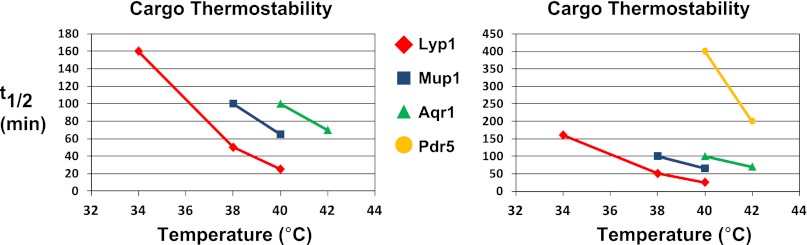
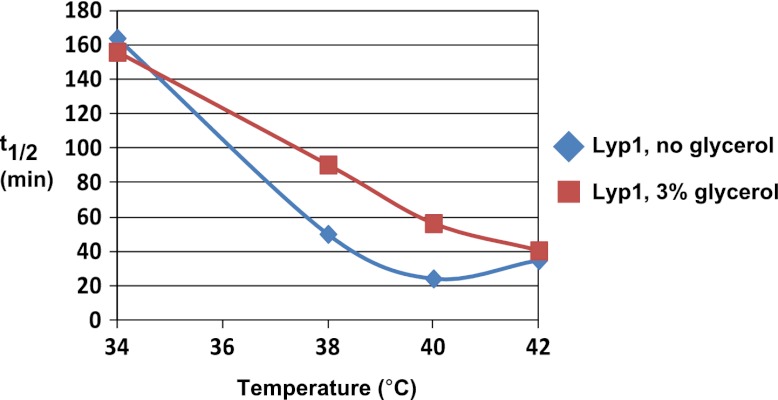
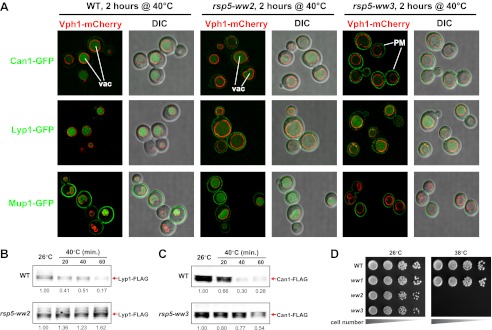
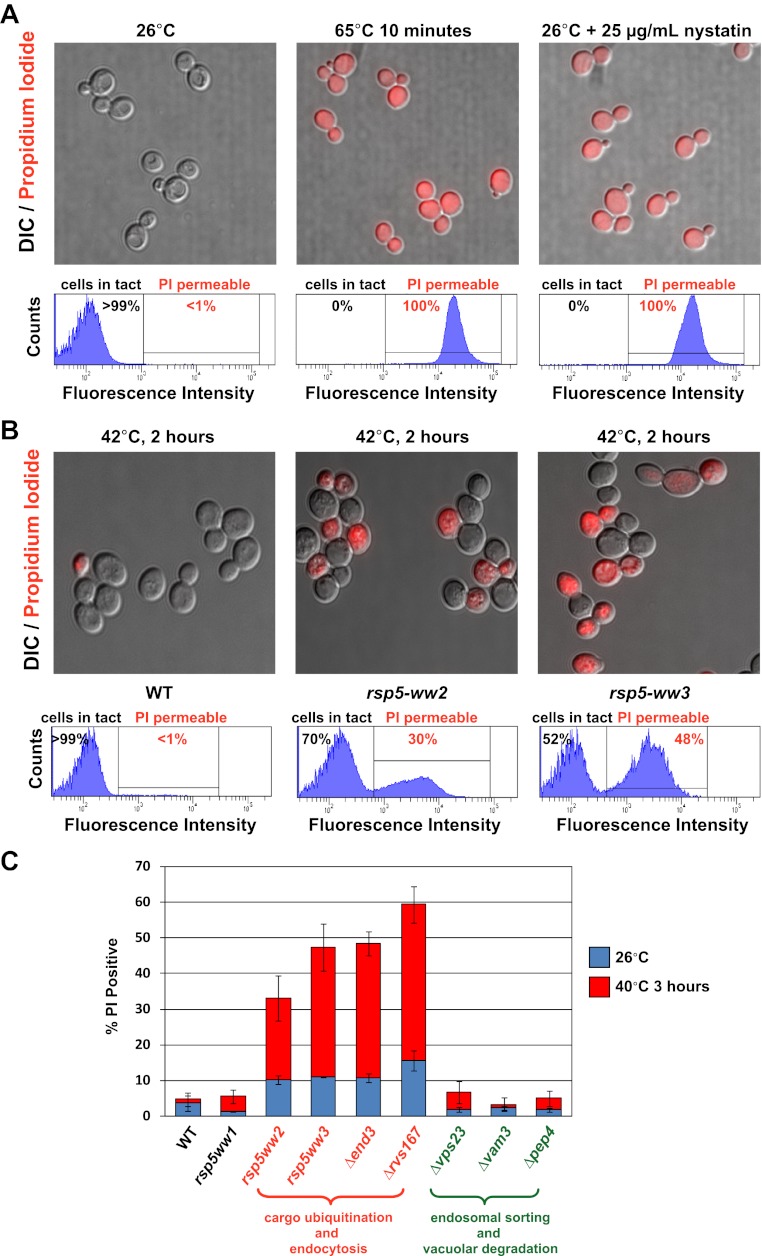
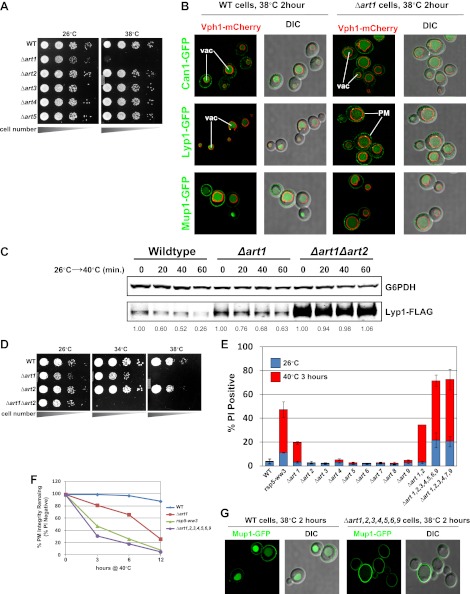
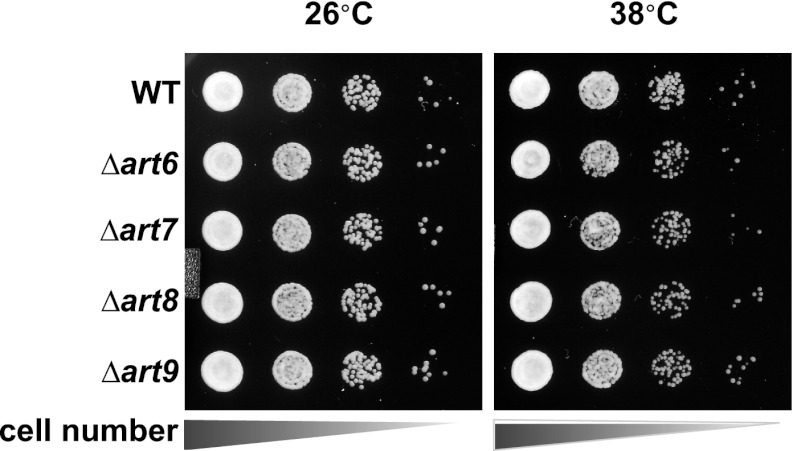
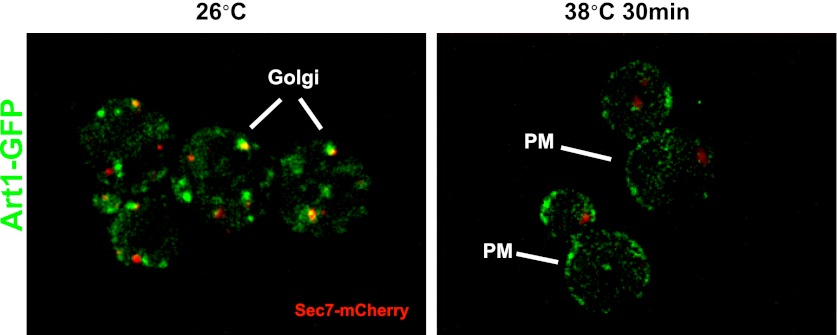
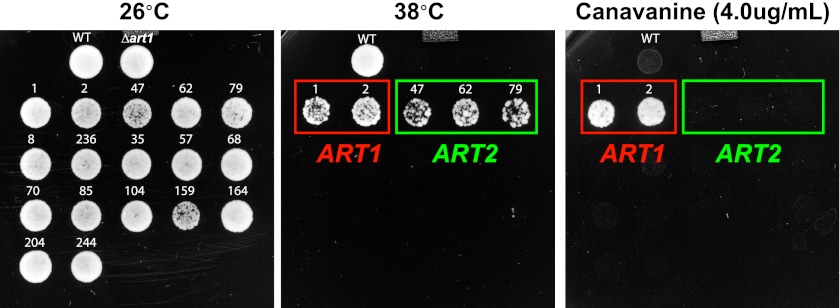
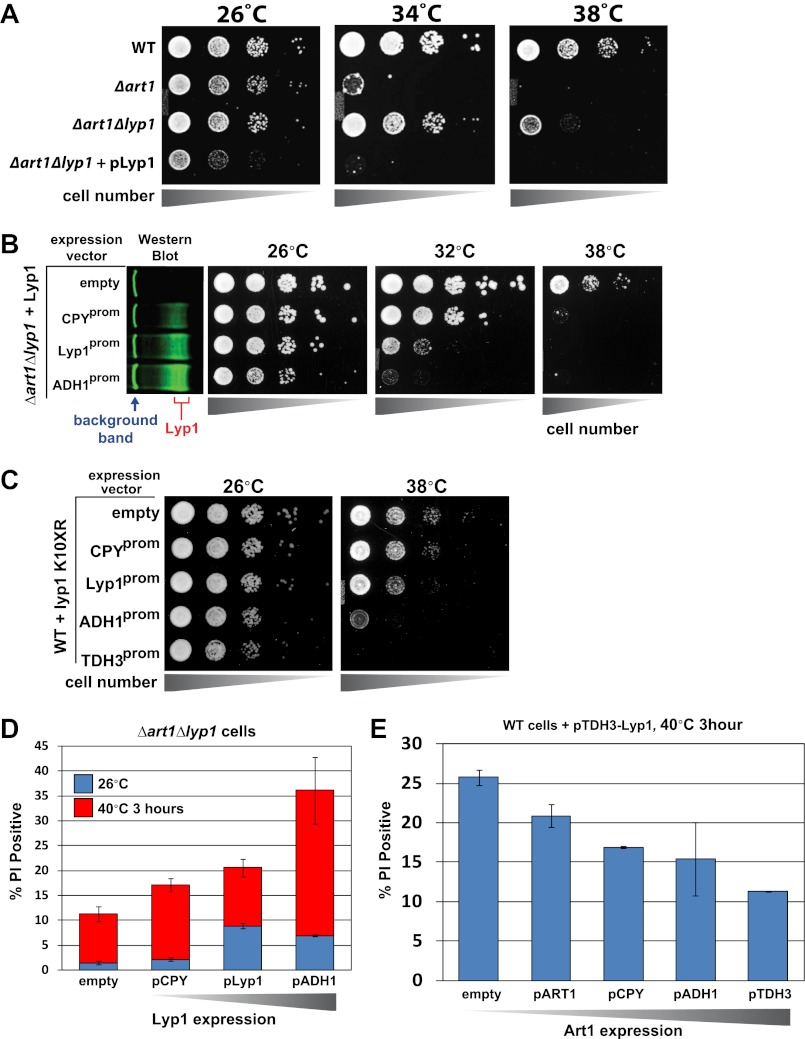
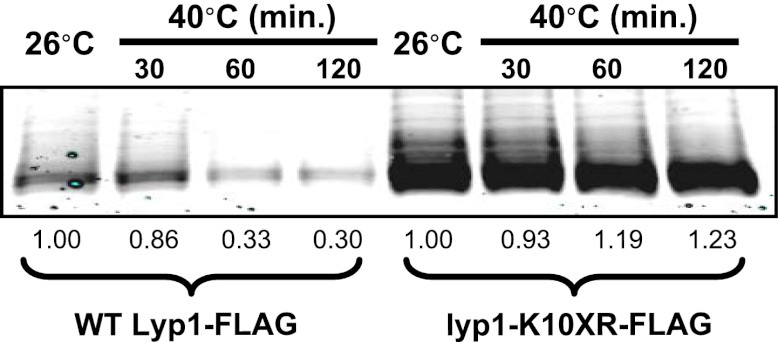
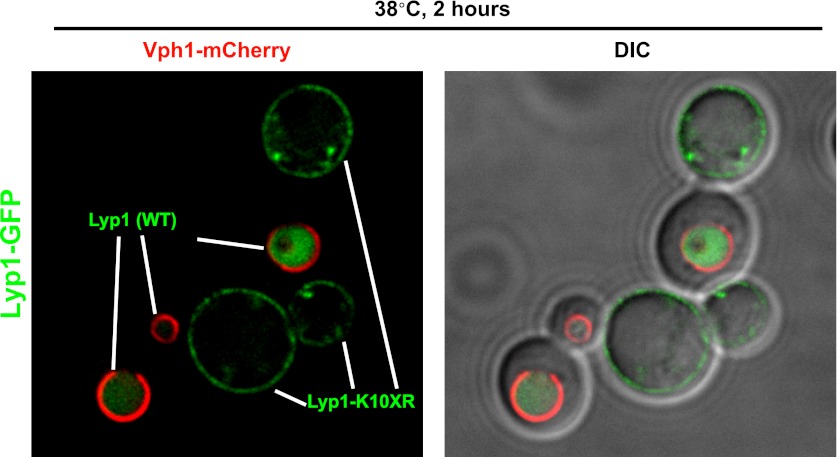
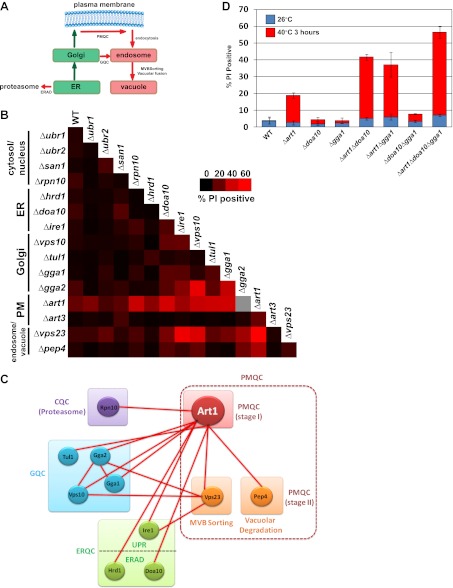
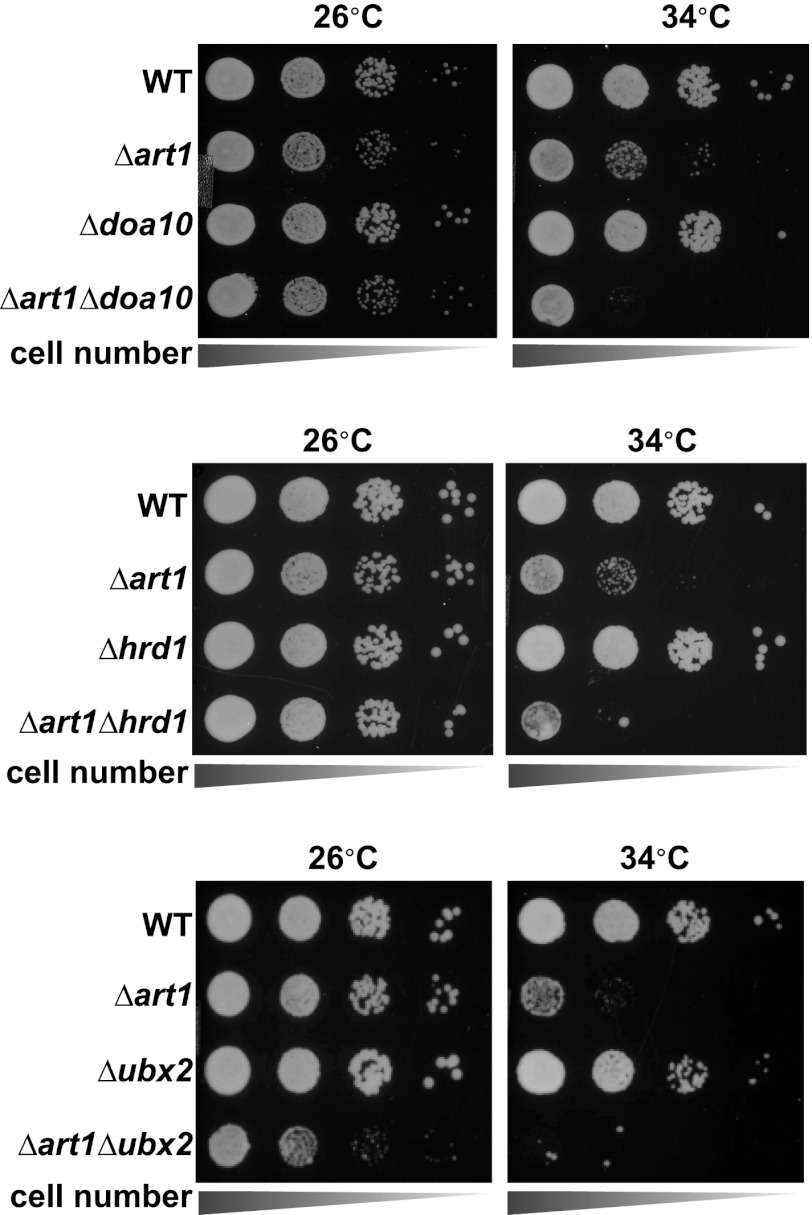
Secretory cargo that cannot fold properly in the ER are selectively targeted for removal by a well-studied ER-associated degradation pathway, or ERAD. In contrast, very little is known about post-ER quality control mechanisms for damaged or misfolded integral membrane proteins. Here we describe a quality control function of the Rsp5-ART ubiquitin ligase adaptor network that functions to protect plasma membrane (PM) integrity. Failure to mediate this protective response during heat stress leads to toxic accumulation of misfolded integral membrane proteins at the cell surface, which causes loss of PM integrity and cell death. Thus, the Rsp5-ART network comprises a PM quality control system that works together with sequential quality control pathways in the ER and Golgi to (i) target the degradation of proteins that have exceeded their functional lifetime due to damage and/or misfolding and (ii) limit the toxic accumulation of specific proteins at the cell surface during proteotoxic stress. DOI:http://dx.doi.org/10.7554/eLife.00459.001.
在内质网(ER)中无法正确折叠的分泌性货物会通过一条经过充分研究的内质网相关降解途径(即ERAD)被选择性地靶向清除。相比之下,对于受损或错误折叠的整合膜蛋白的内质网后质量控制机制,我们了解得非常少。在此,我们描述了Rsp5-ART泛素连接酶衔接子网络的一种质量控制功能,该功能旨在保护质膜(PM)的完整性。在热应激期间未能介导这种保护反应会导致错误折叠的整合膜蛋白在细胞表面毒性积累,进而导致质膜完整性丧失和细胞死亡。因此,Rsp5-ART网络构成了一个质膜质量控制系统,该系统与内质网和高尔基体中的顺序质量控制途径协同作用,以(i)靶向降解因损伤和/或错误折叠而超过其功能寿命的蛋白质,以及(ii)在蛋白毒性应激期间限制特定蛋白质在细胞表面的毒性积累。DOI:http://dx.doi.org/10.7554/eLife.00459.001 。