Vercoe Reuben B, Chang James T, Dy Ron L, Taylor Corinda, Gristwood Tamzin, Clulow James S, Richter Corinna, Przybilski Rita, Pitman Andrew R, Fineran Peter C
Department of Microbiology and Immunology, University of Otago, Dunedin, New Zealand.
PLoS Genet. 2013 Apr;9(4):e1003454. doi: 10.1371/journal.pgen.1003454. Epub 2013 Apr 18.
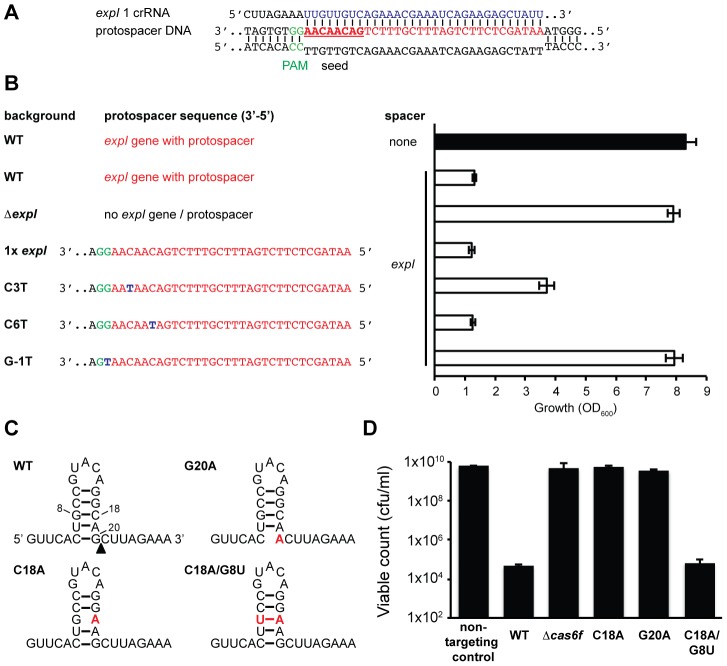
In prokaryotes, clustered regularly interspaced short palindromic repeats (CRISPRs) and their associated (Cas) proteins constitute a defence system against bacteriophages and plasmids. CRISPR/Cas systems acquire short spacer sequences from foreign genetic elements and incorporate these into their CRISPR arrays, generating a memory of past invaders. Defence is provided by short non-coding RNAs that guide Cas proteins to cleave complementary nucleic acids. While most spacers are acquired from phages and plasmids, there are examples of spacers that match genes elsewhere in the host bacterial chromosome. In Pectobacterium atrosepticum the type I-F CRISPR/Cas system has acquired a self-complementary spacer that perfectly matches a protospacer target in a horizontally acquired island (HAI2) involved in plant pathogenicity. Given the paucity of experimental data about CRISPR/Cas-mediated chromosomal targeting, we examined this process by developing a tightly controlled system. Chromosomal targeting was highly toxic via targeting of DNA and resulted in growth inhibition and cellular filamentation. The toxic phenotype was avoided by mutations in the cas operon, the CRISPR repeats, the protospacer target, and protospacer-adjacent motif (PAM) beside the target. Indeed, the natural self-targeting spacer was non-toxic due to a single nucleotide mutation adjacent to the target in the PAM sequence. Furthermore, we show that chromosomal targeting can result in large-scale genomic alterations, including the remodelling or deletion of entire pre-existing pathogenicity islands. These features can be engineered for the targeted deletion of large regions of bacterial chromosomes. In conclusion, in DNA-targeting CRISPR/Cas systems, chromosomal interference is deleterious by causing DNA damage and providing a strong selective pressure for genome alterations, which may have consequences for bacterial evolution and pathogenicity.
在原核生物中,成簇规律间隔短回文重复序列(CRISPRs)及其相关(Cas)蛋白构成了一种抵御噬菌体和质粒的防御系统。CRISPR/Cas系统从外来遗传元件获取短间隔序列,并将这些序列整合到其CRISPR阵列中,形成对过去入侵者的记忆。防御由短非编码RNA提供,这些RNA引导Cas蛋白切割互补核酸。虽然大多数间隔序列是从噬菌体和质粒获取的,但也有一些间隔序列与宿主细菌染色体其他位置的基因匹配的例子。在黑胫果胶杆菌中,I-F型CRISPR/Cas系统获得了一个自我互补的间隔序列,该序列与参与植物致病性的水平获得岛(HAI2)中的原间隔序列靶点完美匹配。鉴于关于CRISPR/Cas介导的染色体靶向的实验数据匮乏,我们通过开发一个严格控制的系统来研究这一过程。通过靶向DNA进行染色体靶向具有高度毒性,会导致生长抑制和细胞丝状化。通过对cas操纵子、CRISPR重复序列、原间隔序列靶点以及靶点旁边的原间隔序列相邻基序(PAM)进行突变,可以避免毒性表型。事实上,天然的自我靶向间隔序列由于PAM序列中靶点旁边的单个核苷酸突变而无毒。此外,我们表明染色体靶向可导致大规模基因组改变,包括整个预先存在的致病岛的重塑或缺失。这些特性可用于对细菌染色体的大片段进行靶向缺失。总之,在靶向DNA的CRISPR/Cas系统中,染色体干扰通过造成DNA损伤并为基因组改变提供强大的选择压力而具有有害性,这可能对细菌进化和致病性产生影响。