Institute of Ecology and Evolution, University of Oregon, Eugene, Oregon, United States of America.
Biochemistry Department, Emory University School of Medicine, Atlanta, Georgia, United States of America.
PLoS Genet. 2014 Jan;10(1):e1004058. doi: 10.1371/journal.pgen.1004058. Epub 2014 Jan 9.
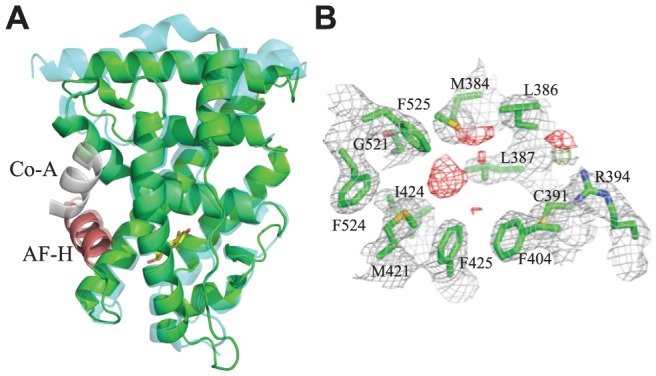
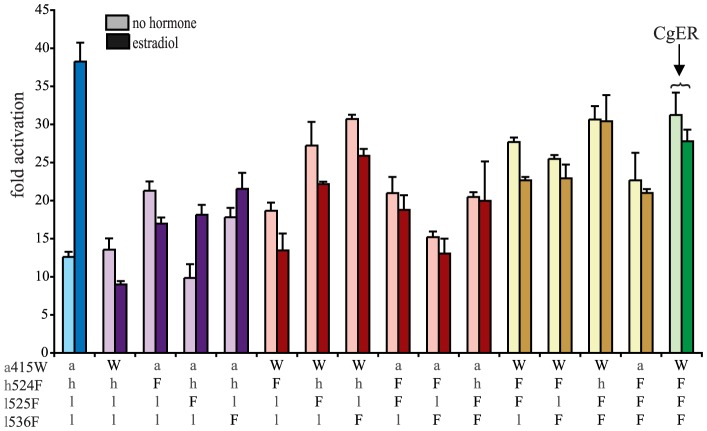
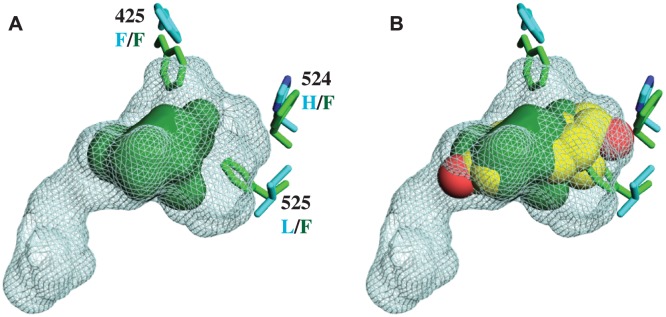
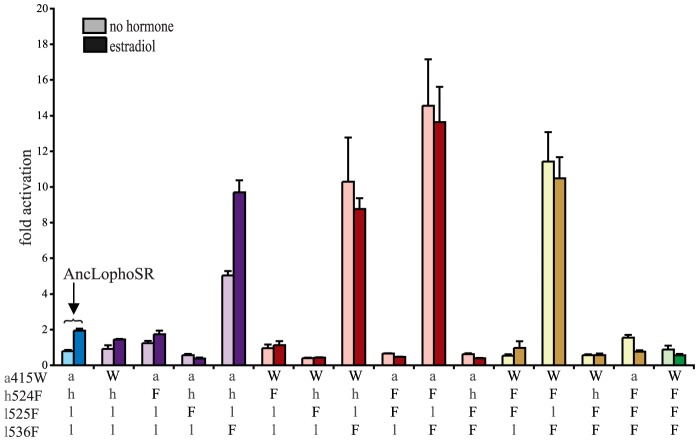
An important goal in molecular evolution is to understand the genetic and physical mechanisms by which protein functions evolve and, in turn, to characterize how a protein's physical architecture influences its evolution. Here we dissect the mechanisms for an evolutionary shift in function in the mollusk ortholog of the steroid hormone receptors (SRs), a family of biologically essential transcription factors. In vertebrates, the activity of SRs allosterically depends on binding a hormonal ligand; in mollusks, however, the SR ortholog (called ER, because of high sequence similarity to vertebrate estrogen receptors) activates transcription in the absence of ligand and does not respond to steroid hormones. To understand how this shift in regulation evolved, we combined evolutionary, structural, and functional analyses. We first determined the X-ray crystal structure of the ER of the Pacific oyster Crassostrea gigas (CgER), and found that its ligand pocket is filled with bulky residues that prevent ligand occupancy. To understand the genetic basis for the evolution of mollusk ERs' unique functions, we resurrected an ancient SR progenitor and characterized the effect of historical amino acid replacements on its functions. We found that reintroducing just two ancient replacements from the lineage leading to mollusk ERs recapitulates the evolution of full constitutive activity and the loss of ligand activation. These substitutions stabilize interactions among key helices, causing the allosteric switch to become "stuck" in the active conformation and making activation independent of ligand binding. Subsequent changes filled the ligand pocket without further affecting activity; by degrading the allosteric switch, these substitutions vestigialized elements of the protein's architecture required for ligand regulation and made reversal to the ancestral function more complex. These findings show how the physical architecture of allostery enabled a few large-effect mutations to trigger a profound evolutionary change in the protein's function and shaped the genetics of evolutionary reversibility.
分子进化的一个重要目标是理解蛋白质功能进化的遗传和物理机制,并进而描述蛋白质的物理结构如何影响其进化。在这里,我们剖析了软体动物甾体激素受体(SRs)同源物功能进化转变的机制,SRs 是一类具有重要生物学功能的转录因子。在脊椎动物中,SR 的活性通过与激素配体结合而变构依赖;然而,在软体动物中,SR 同源物(由于与脊椎动物雌激素受体具有高度序列相似性,称为 ER)在没有配体的情况下激活转录,并且不响应类固醇激素。为了理解这种调控转变是如何进化的,我们结合了进化、结构和功能分析。我们首先确定了太平洋牡蛎(Crassostrea gigas)ER 的 X 射线晶体结构,并发现其配体口袋充满了阻止配体占据的大体积残基。为了理解软体动物 ER 独特功能进化的遗传基础,我们复活了一个古老的 SR 前体,并研究了历史氨基酸替换对其功能的影响。我们发现,仅从导致软体动物 ER 的谱系中引入两个古老的替换就可以再现完全组成型活性的进化和配体激活的丧失。这些取代稳定了关键螺旋之间的相互作用,导致变构开关“卡在”活性构象中,使激活独立于配体结合。随后的变化填满了配体口袋,而没有进一步影响活性;通过降解变构开关,这些取代使蛋白结构中用于配体调节的遗迹元素退化,并使向祖先功能的逆转变得更加复杂。这些发现表明,变构的物理结构如何使少数大效应突变触发蛋白功能的深刻进化变化,并塑造了进化可逆性的遗传学。