Zhang Hesheng, Gao Lu, Anandhakumar Jayamani, Gross David S
Department of Biochemistry and Molecular Biology, Louisiana State University Health Sciences Center, Shreveport, Louisiana, United States of America.
PLoS Genet. 2014 Apr 10;10(4):e1004202. doi: 10.1371/journal.pgen.1004202. eCollection 2014 Apr.
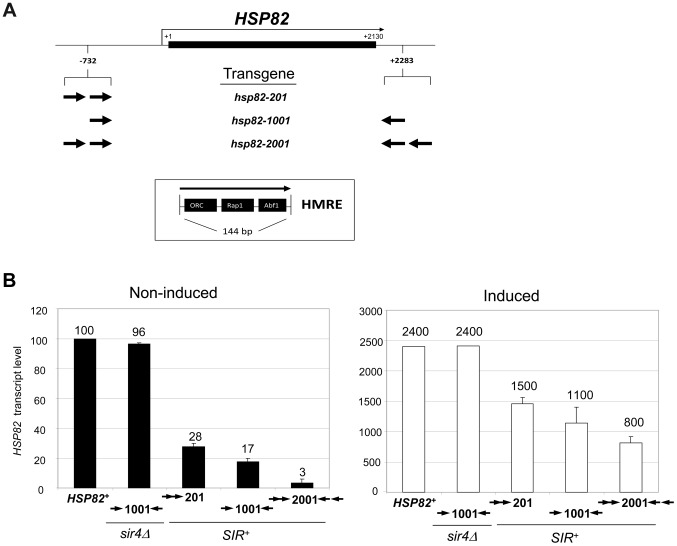
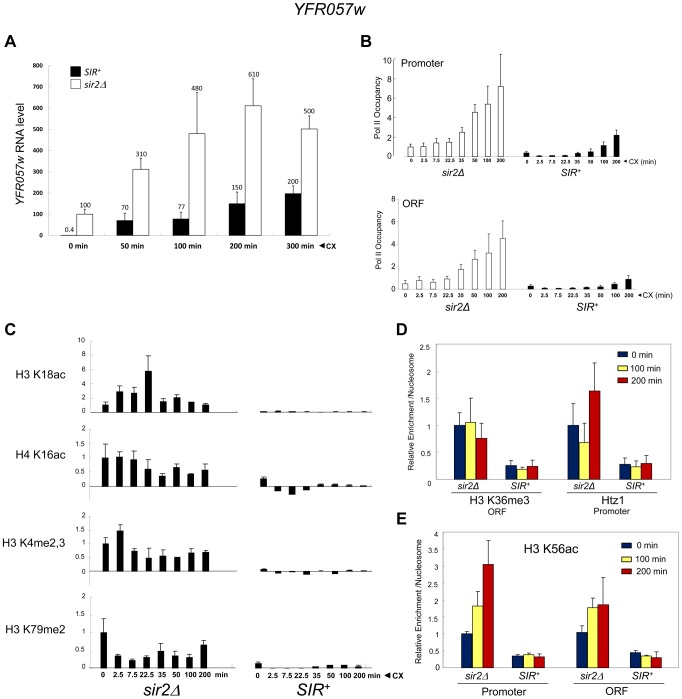
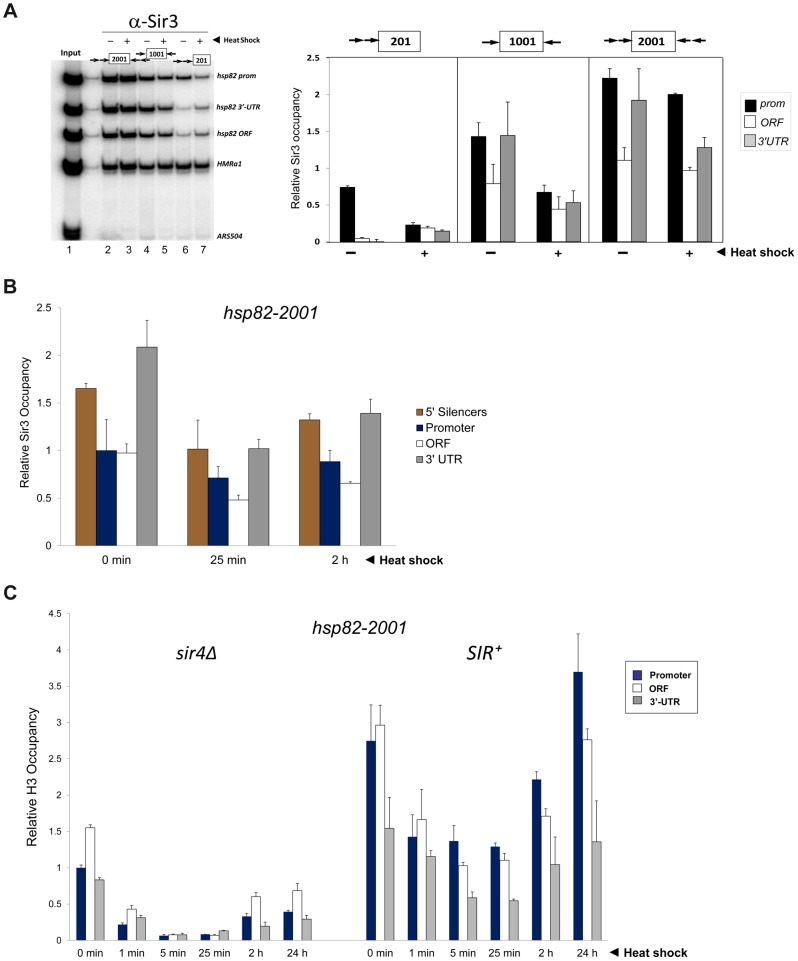
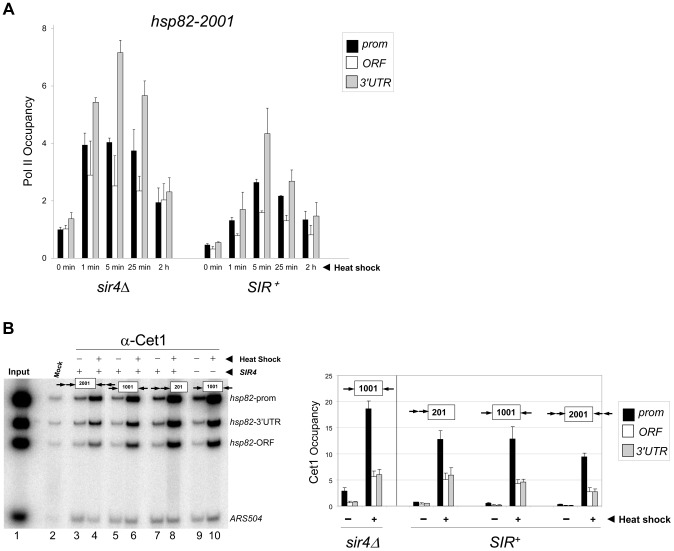
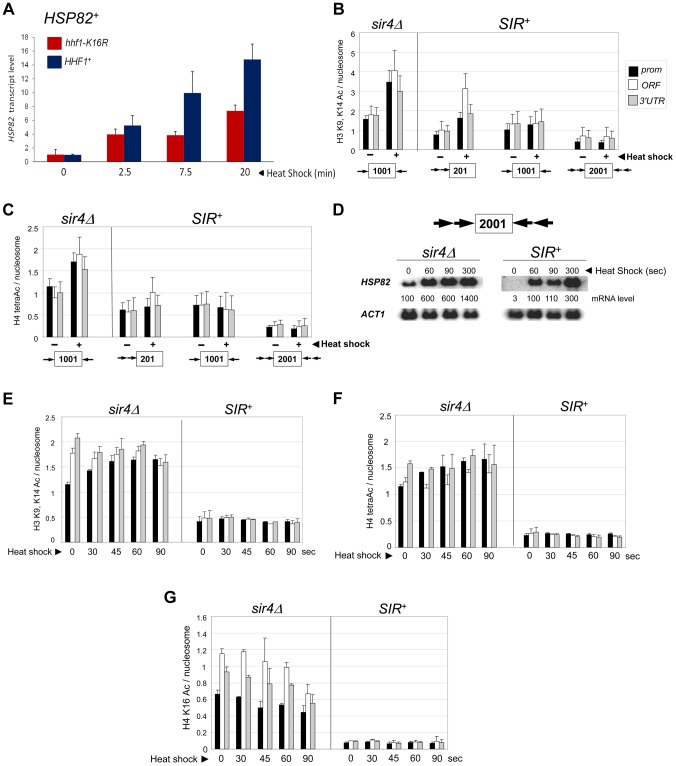
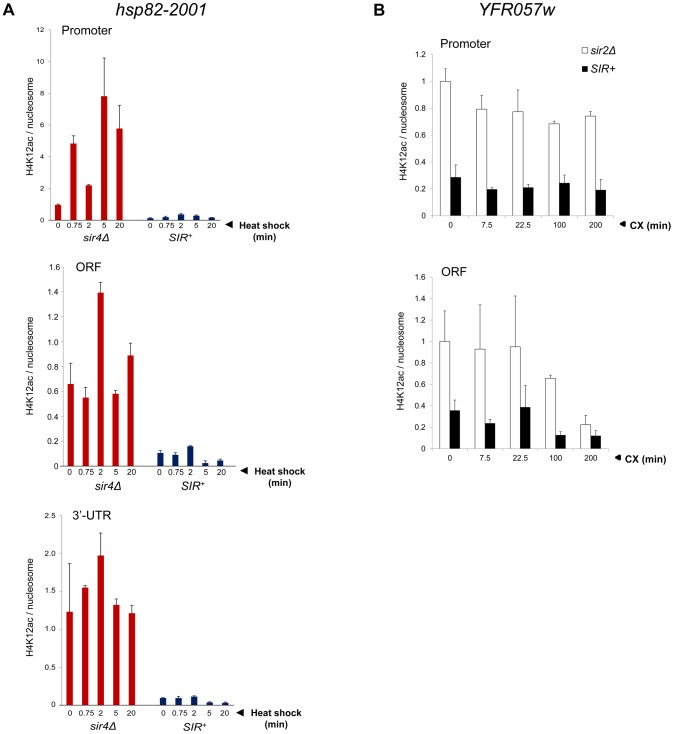
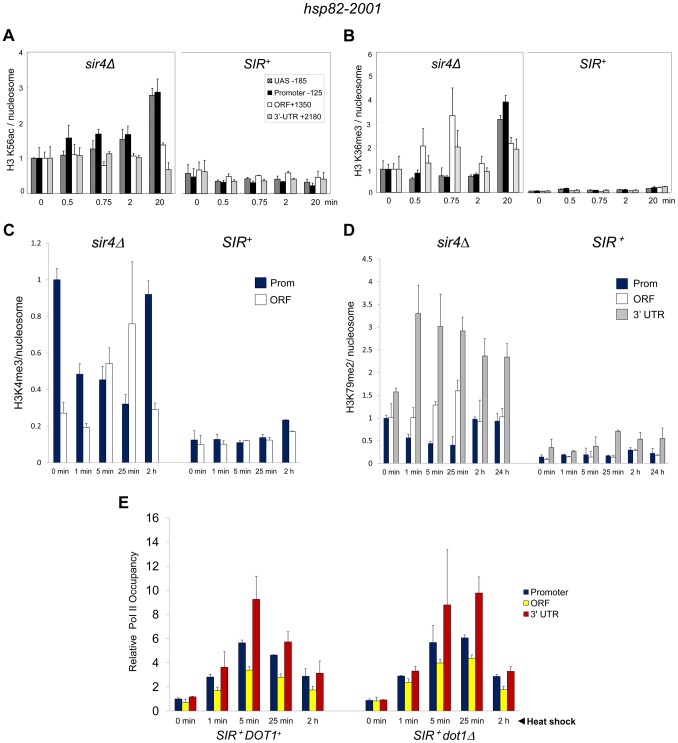
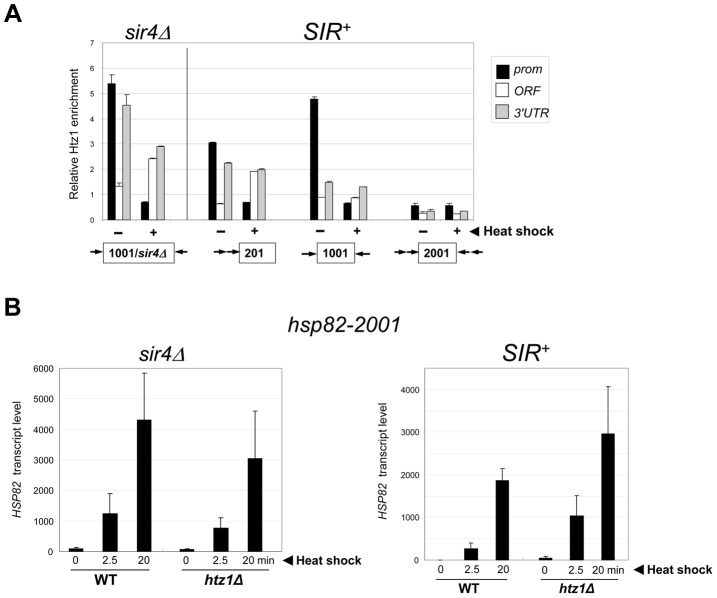
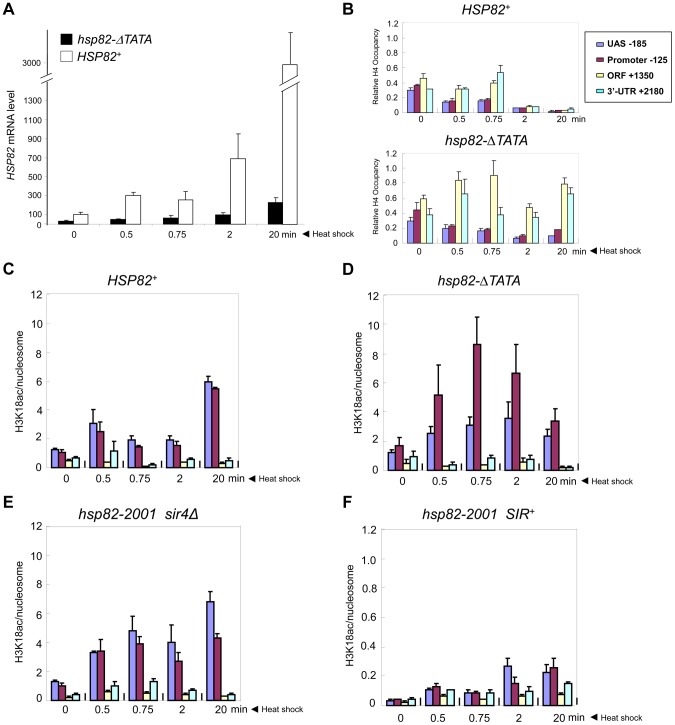
It is widely accepted that transcriptional regulation of eukaryotic genes is intimately coupled to covalent modifications of the underlying chromatin template, and in certain cases the functional consequences of these modifications have been characterized. Here we present evidence that gene activation in the silent heterochromatin of the yeast Saccharomyces cerevisiae can occur in the context of little, if any, covalent histone modification. Using a SIR-regulated heat shock-inducible transgene, hsp82-2001, and a natural drug-inducible subtelomeric gene, YFR057w, as models we demonstrate that substantial transcriptional induction (>200-fold) can occur in the context of restricted histone loss and negligible levels of H3K4 trimethylation, H3K36 trimethylation and H3K79 dimethylation, modifications commonly linked to transcription initiation and elongation. Heterochromatic gene activation can also occur with minimal H3 and H4 lysine acetylation and without replacement of H2A with the transcription-linked variant H2A.Z. Importantly, absence of histone modification does not stem from reduced transcriptional output, since hsp82-ΔTATA, a euchromatic promoter mutant lacking a TATA box and with threefold lower induced transcription than heterochromatic hsp82-2001, is strongly hyperacetylated in response to heat shock. Consistent with negligible H3K79 dimethylation, dot1Δ cells lacking H3K79 methylase activity show unimpeded occupancy of RNA polymerase II within activated heterochromatic promoter and coding regions. Our results indicate that large increases in transcription can be observed in the virtual absence of histone modifications often thought necessary for gene activation.
人们普遍认为,真核基因的转录调控与潜在染色质模板的共价修饰密切相关,并且在某些情况下,这些修饰的功能后果已得到表征。在这里,我们提供证据表明,酿酒酵母沉默异染色质中的基因激活可以在很少(如果有的话)共价组蛋白修饰的情况下发生。使用受SIR调控的热休克诱导转基因hsp82 - 2001和天然药物诱导的亚端粒基因YFR057w作为模型,我们证明在组蛋白损失受限以及H3K4三甲基化、H3K36三甲基化和H3K79二甲基化水平可忽略不计的情况下,可以发生大量的转录诱导(>200倍),这些修饰通常与转录起始和延伸相关。异染色质基因激活也可以在H3和H4赖氨酸乙酰化水平极低且没有用与转录相关的变体H2A.Z替代H2A的情况下发生。重要的是,组蛋白修饰的缺失并非源于转录输出减少,因为hsp82 - ΔTATA是一个常染色质启动子突变体,缺乏TATA框,其诱导转录比异染色质的hsp82 - 2001低三倍,但在热休克反应中强烈超乙酰化。与可忽略不计的H3K79二甲基化一致,缺乏H3K79甲基化酶活性的dot1Δ细胞在激活的异染色质启动子和编码区域内RNA聚合酶II的占据不受阻碍。我们的结果表明,在通常认为基因激活所必需的组蛋白修饰几乎不存在的情况下,可以观察到转录的大幅增加。