Ye Z P, Baylor N W, Wagner R R
Department of Microbiology, University of Virginia Medical School, Charlottesville 22908.
J Virol. 1989 Sep;63(9):3586-94. doi: 10.1128/JVI.63.9.3586-3594.1989.
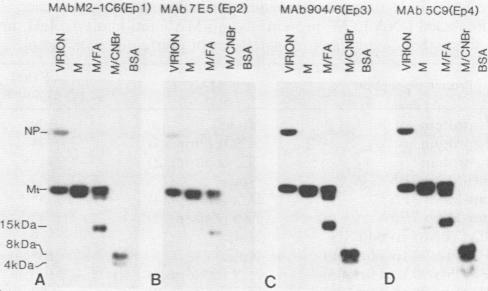
We have undertaken by biochemical and immunological experiments to locate the region of the matrix (M1) protein responsible for down-regulating endogenous transcription of A/WSN/33 influenza virus. A more refined map of the antigenic determinants of the M1 protein was obtained by binding of epitope-specific monoclonal antibodies (MAbs) to chemically cleaved fragments. Epitope 2-specific MAb 289/4 and MAb 7E5 reverse transcription inhibition by M1 protein and react with a 4-kilodalton cyanogen bromide fragment extending from amino acid Gly-129 to Gln-164. Anti-idiotype serum immunoglobulin G prepared in rabbits immunized with MAb 289/4 or MAb 7E5 mimicked the action of M1 protein by inhibiting transcription in vitro of influenza virus ribonucleoprotein cores. This transcription-inhibition activity of anti-MAb 7E5 immunoglobulin G and anti-MAb 289/4 immunoglobulin G could be reversed by MAb 7E5 and MAb 289/4 or could be removed by MAb 7E5-Sepharose affinity chromatography. Transcription of influenza virus ribonucleoprotein was inhibited by one of three synthetic oligopeptides, a nonodecapeptide SP3 with an amino acid sequence corresponding to Pro-90 through Thr-108 of the M1 protein. Of all the structural proteins of influenza virus, only NP and M1 showed strong affinity for binding viral RNA or other extraneous RNAs. The 4-kilodalton cyanogen bromide peptide (Gly-129 to Gln-164), exhibited marked affinity for viral RNA, the binding of which was blocked by epitope 2-specific MAb 7E5 but not by MAbs directed to three other epitopes. Viral RNA also bound strongly to the nonodecapeptide SP3 and rather less well to anti-idiotype anti-MAb 7E5; these latter viral RNA-binding reactions were only slightly blocked by preincubation of anti-MAb 7E5 or SP3 with MAb 7E5. These experiments suggest the presence of at least two RNA-binding sites, which also serve as transcription-inhibition sites, centered around amino acid sequences 80 through 109 (epitope 4?) and 129 through 164 (epitope 2) of the 252 amino acid M1 protein of A/WSN/33 influenza virus. A hydropathy plot of the M1 protein calculated by free-energy transfer suggests that the two hydrophilic transcription-inhibition RNA-binding domains are brought into close proximity by an alpha-helix-forming intervening hydrophobic domain.
我们通过生化和免疫学实验来确定基质(M1)蛋白中负责下调A/WSN/33流感病毒内源性转录的区域。通过表位特异性单克隆抗体(MAb)与化学裂解片段的结合,获得了更精细的M1蛋白抗原决定簇图谱。表位2特异性单克隆抗体289/4和单克隆抗体7E5可逆转M1蛋白的逆转录抑制作用,并与一个从氨基酸Gly-129延伸至Gln-164的4千道尔顿溴化氰片段发生反应。在用单克隆抗体289/4或单克隆抗体7E5免疫的兔中制备的抗独特型血清免疫球蛋白G,通过抑制流感病毒核糖核蛋白核心的体外转录来模拟M1蛋白的作用。抗单克隆抗体7E5免疫球蛋白G和抗单克隆抗体289/4免疫球蛋白G的这种转录抑制活性可被单克隆抗体7E5和单克隆抗体289/4逆转,或可通过单克隆抗体7E5-琼脂糖亲和层析去除。流感病毒核糖核蛋白的转录被三种合成寡肽之一抑制,即一种十九肽SP3,其氨基酸序列对应于M1蛋白的Pro-90至Thr-108。在流感病毒的所有结构蛋白中,只有核蛋白(NP)和M1蛋白对结合病毒RNA或其他外来RNA表现出强亲和力。4千道尔顿溴化氰肽(Gly-129至Gln-164)对病毒RNA表现出显著亲和力,其结合被表位2特异性单克隆抗体7E5阻断,但不被针对其他三个表位的单克隆抗体阻断。病毒RNA也与十九肽SP3强烈结合,与抗独特型抗单克隆抗体7E5的结合较弱;后一种病毒RNA结合反应仅被抗单克隆抗体7E5或SP3与单克隆抗体7E5的预孵育轻微阻断。这些实验表明,在A/WSN/33流感病毒252个氨基酸的M1蛋白中,至少存在两个RNA结合位点,它们也作为转录抑制位点,集中在氨基酸序列80至109(表位4?)和129至164(表位2)周围。通过自由能转移计算得到的M1蛋白亲水性图谱表明,两个亲水性转录抑制RNA结合结构域通过一个形成α螺旋的中间疏水性结构域紧密靠近。