Padilla-Parra Sergi, Marin Mariana, Kondo Naoyuki, Melikyan Gregory B
Division of Pediatric Infectious Diseases, Emory University Children's Center, Atlanta, GA 30322, USA.
Retrovirology. 2014 Jun 16;11:47. doi: 10.1186/1742-4690-11-47.
The majority of viruses enter host cells via endocytosis. Current knowledge of viral entry pathways is largely based upon infectivity measurements following genetic and/or pharmacological interventions that disrupt vesicular trafficking and maturation. Imaging of single virus entry in living cells provides a powerful means to delineate viral trafficking pathways and entry sites under physiological conditions.
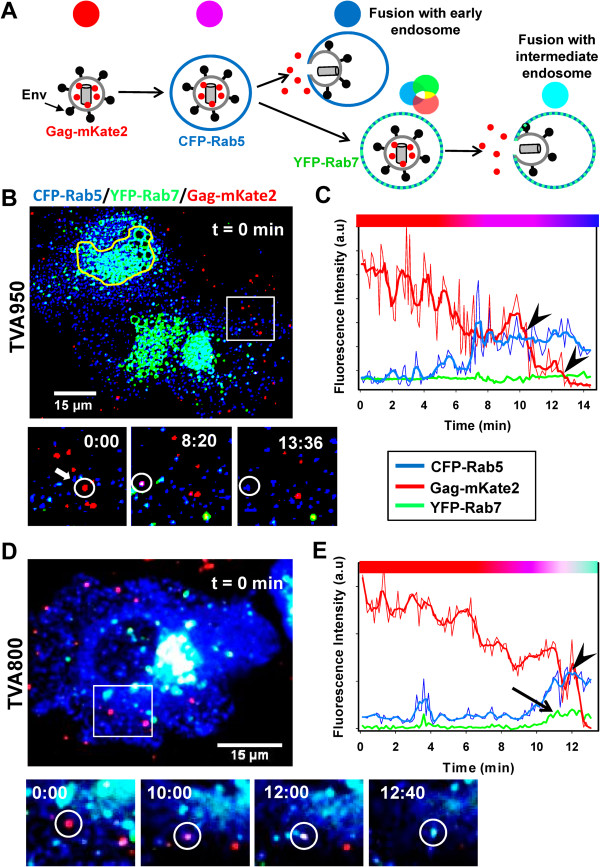
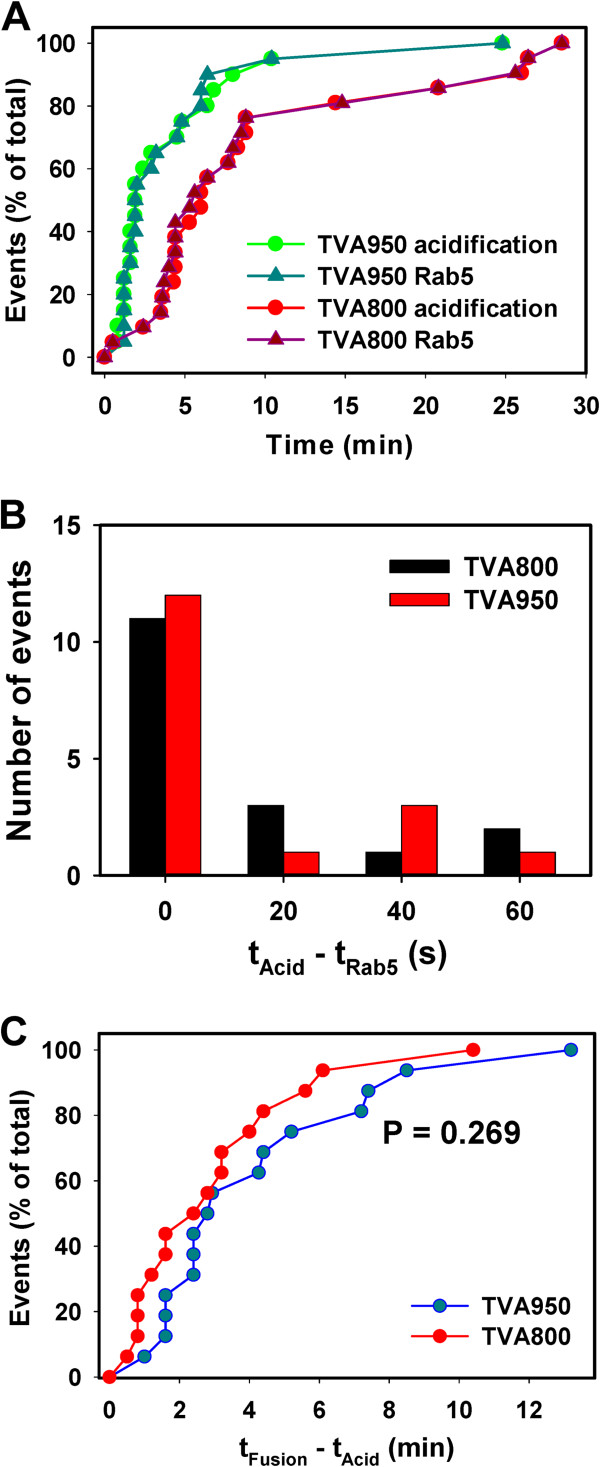
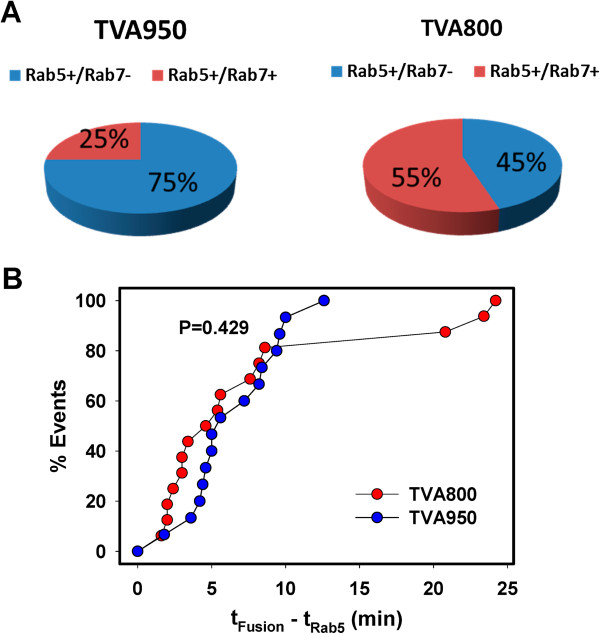
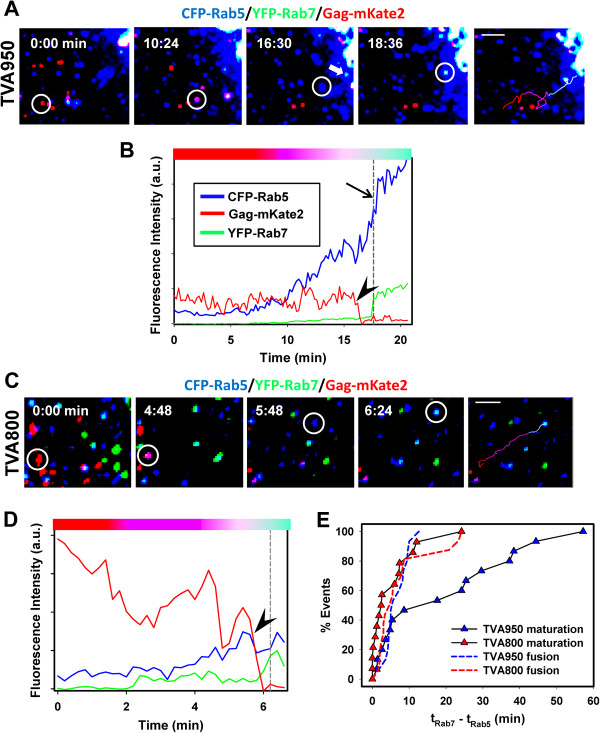
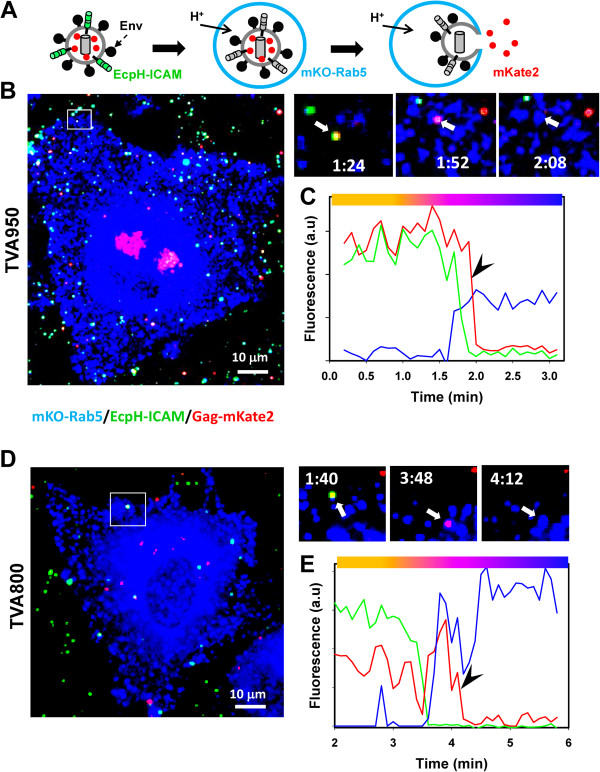
Here, we visualized single avian retrovirus co-trafficking with markers for early (Rab5) and late (Rab7) endosomes, acidification of endosomal lumen and the resulting viral fusion measured by the viral content release into the cytoplasm. Virus-carrying vesicles either merged with the existing Rab5-positive early endosomes or slowly accumulated Rab5. The Rab5 recruitment to virus-carrying endosomes correlated with acidification of their lumen. Viral fusion occurred either in early (Rab5-positive) or intermediate (Rab5- and Rab7-positive) compartments. Interestingly, different isoforms of the cognate receptor directed virus entry from distinct endosomes. In cells expressing the transmembrane receptor, viruses preferentially entered and fused with slowly maturing early endosomes prior to accumulation of Rab7. By comparison, in cells expressing the GPI-anchored receptor, viruses entered both slowly and quickly maturing endosomes and fused with early (Rab5-positive) and intermediate (Rab5- and Rab7-positive) compartments.
Since the rate of low pH-triggered fusion was independent of the receptor isoform, we concluded that the sites of virus entry are determined by the kinetic competition between endosome maturation and viral fusion. Our findings demonstrate the ability of this retrovirus to enter cells via alternative endocytic pathways and establish infection by releasing its content from distinct endosomal compartments.
大多数病毒通过内吞作用进入宿主细胞。目前关于病毒进入途径的知识主要基于对遗传和/或药理学干预后感染性的测量,这些干预会破坏囊泡运输和成熟过程。对活细胞中单个病毒进入过程的成像为描绘生理条件下病毒运输途径和进入位点提供了有力手段。
在这里,我们观察到单个禽逆转录病毒与早期(Rab5)和晚期(Rab7)内体标记物共同运输,测量了内体腔的酸化以及通过病毒内容物释放到细胞质中所导致的病毒融合。携带病毒的囊泡要么与现有的Rab5阳性早期内体融合,要么缓慢积累Rab5。Rab5募集到携带病毒的内体与其腔的酸化相关。病毒融合发生在早期(Rab5阳性)或中间(Rab5和Rab7阳性)区室。有趣的是,同源受体的不同异构体引导病毒从不同的内体进入。在表达跨膜受体的细胞中,病毒在Rab7积累之前优先进入并与缓慢成熟的早期内体融合。相比之下,在表达糖基磷脂酰肌醇锚定受体的细胞中,病毒进入缓慢和快速成熟的内体,并与早期(Rab5阳性)和中间(Rab5和Rab7阳性)区室融合。
由于低pH触发的融合速率与受体异构体无关,我们得出结论,病毒进入位点由内体成熟和病毒融合之间的动力学竞争决定。我们的研究结果表明这种逆转录病毒能够通过替代内吞途径进入细胞,并通过从不同的内体区室释放其内容物来建立感染。