Tsanakas Georgios F, Manioudaki Maria E, Economou Athanasios S, Kalaitzis Panagiotis
Department of Horticultural Genetics & Biotechnology, Mediterranean Agronomic Institute of Chania (MAICh), Crete, Greece.
BMC Genomics. 2014 Jul 4;15(1):554. doi: 10.1186/1471-2164-15-554.
The petal senescence of ethylene insensitive species has not been investigated thoroughly while little is known about the temporal and tissue specific expression patterns of transcription factors (TFs) in this developmental process. Even less is known on flower senescence of the ornamental pot plant Gardenia jasminoides, a non climacteric flower with significant commercial value.
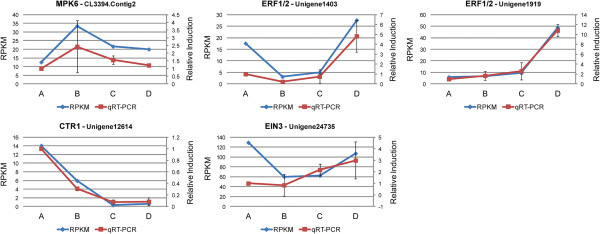
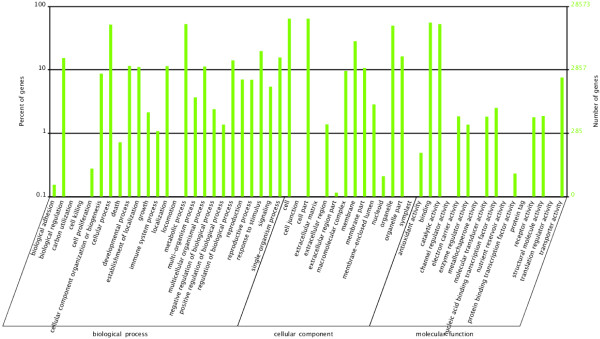
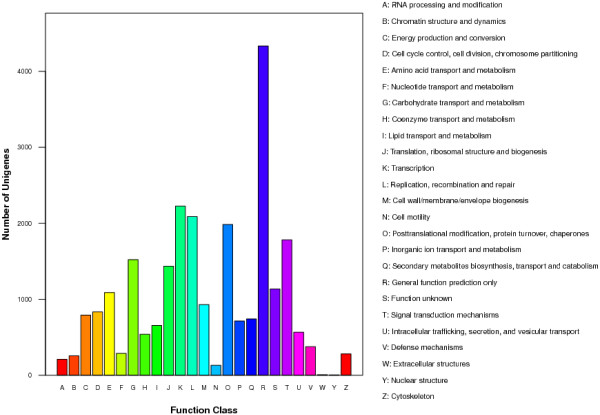
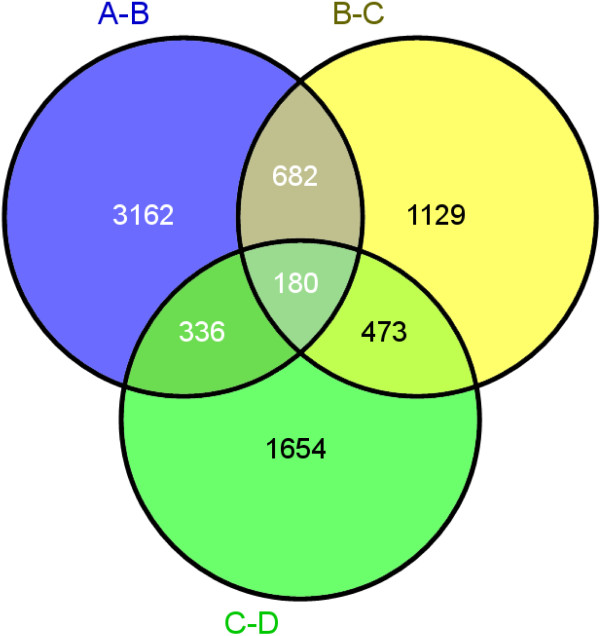
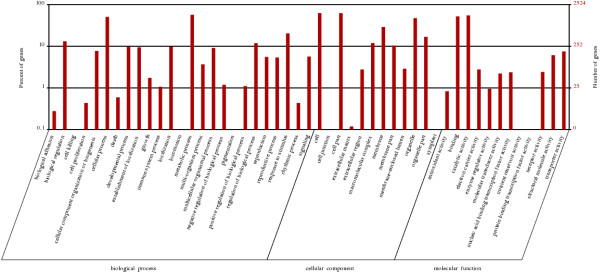
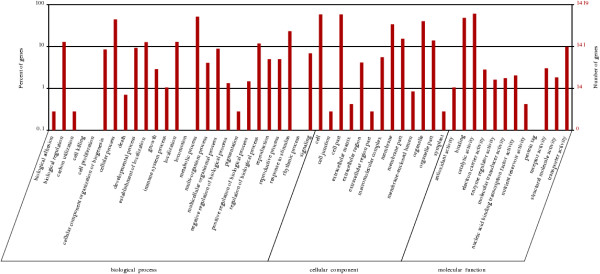
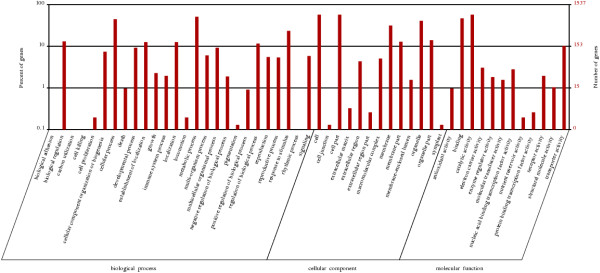
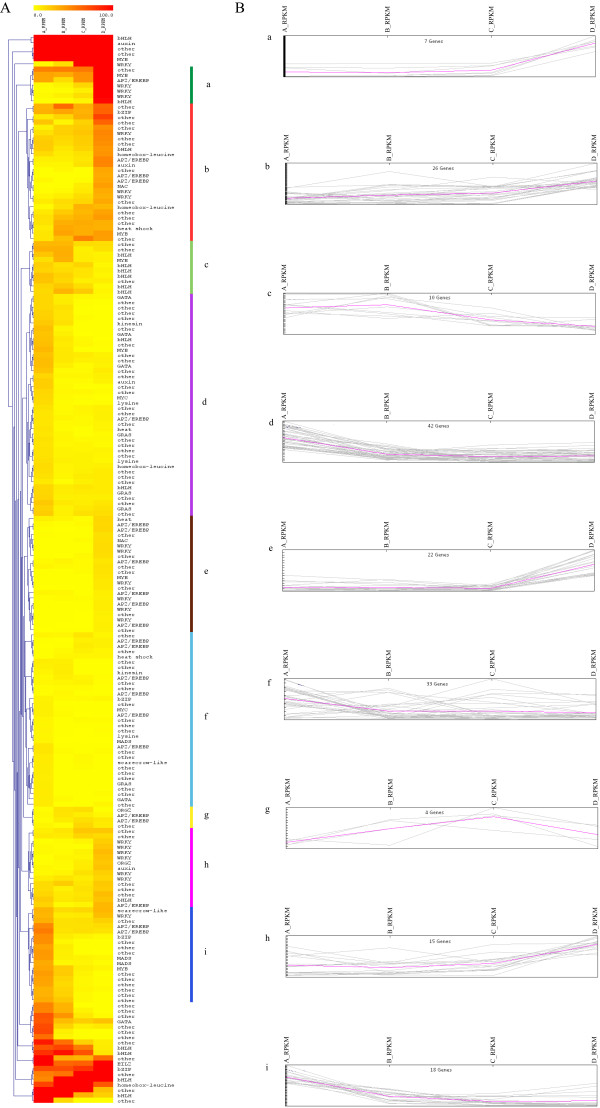
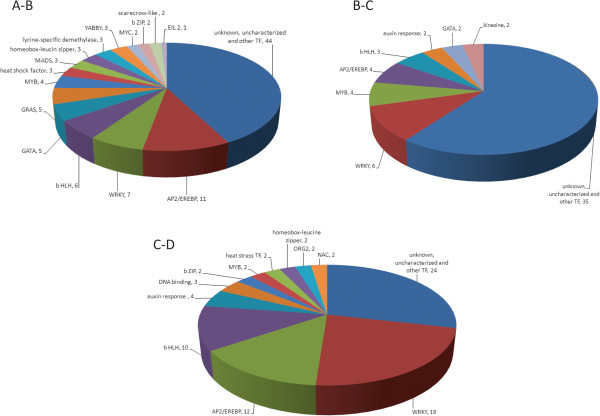
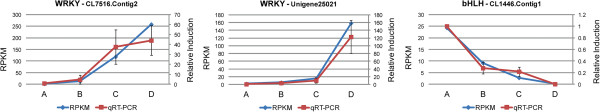
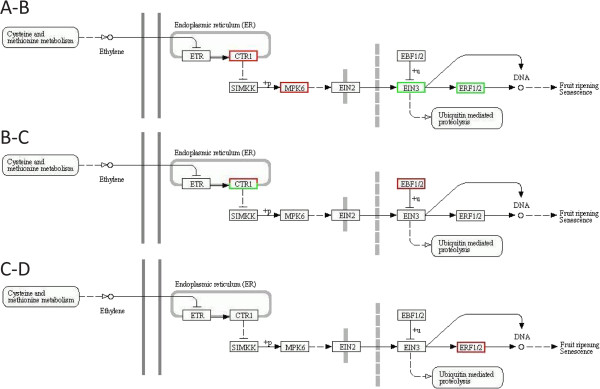
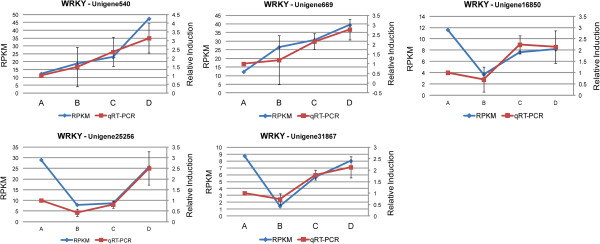
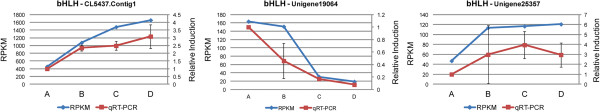
We initiated a de novo transcriptome study to investigate the petal senescence in four developmental stages of cut gardenia flowers considering that the visible symptoms of senescence appear within 4 days of flower opening. De novo assembly of transcriptome sequencing resulted in 102,263 contigs with mean length of 360 nucleotides that generated 57,503 unigenes. These were further clustered into 20,970 clusters and 36,533 singletons. The comparison of the consecutive developmental stages resulted in 180 common, differentially expressed unigenes. A large number of Simple Sequence Repeats were also identified comprising a large number of dinucleotides and trinucleotides. The prevailing families of differentially expressed TFs comprise the AP2/EREBP, WRKY and the bHLH. There are 81 differentially expressed TFs when the symptoms of flower senescence become visible with the most prevailing being the WRKY family with 19 unigenes. No other WRKY TFs had been identified up to now in petal senescence of ethylene insensitive species. A large number of differentially expressed genes were identified at the initiation of visible symptoms of senescence compared to the open flower stage indicating a significant shift in the expression profiles which might be coordinated by up-regulated and/or down-regulated TFs. The expression of 16 genes that belong to the TF families of WRKY, bHLH and the ethylene sensing pathway was validated using qRT--PCR.
This de novo transcriptome analysis resulted in the identification of TFs with specific temporal expression patterns such as two WRKYs and one bHLH, which might play the role of senescence progression regulators. Further research is required to investigate their role in gardenia flowers in order to develop tools to delay petal senescence.
乙烯不敏感型物种的花瓣衰老尚未得到充分研究,对于转录因子(TFs)在这一发育过程中的时空表达模式也知之甚少。关于具有重要商业价值的非跃变型观赏盆栽植物栀子的花衰老,了解得更少。
考虑到切花栀子在开花后4天内出现衰老的可见症状,我们开展了一项从头转录组研究,以调查切花栀子四个发育阶段的花瓣衰老情况。转录组测序的从头组装产生了102,263个重叠群,平均长度为360个核苷酸,生成了57,503个单基因。这些进一步聚类为20,970个簇和36,533个单拷贝。连续发育阶段的比较产生了180个共同的、差异表达的单基因。还鉴定出大量简单序列重复,其中包含大量二核苷酸和三核苷酸。差异表达的TFs的主要家族包括AP2/EREBP、WRKY和bHLH。当花衰老症状可见时,有81个差异表达的TFs,其中最主要的是WRKY家族,有19个单基因。到目前为止,在乙烯不敏感型物种的花瓣衰老中尚未鉴定出其他WRKY TFs。与开放花阶段相比,在衰老可见症状开始时鉴定出大量差异表达基因,表明表达谱发生了显著变化,这可能由上调和/或下调的TFs协调。使用qRT-PCR验证了属于WRKY、bHLH和乙烯感知途径TF家族的16个基因的表达。
这项从头转录组分析鉴定出具有特定时间表达模式的TFs,如两个WRKY和一个bHLH,它们可能起到衰老进程调节因子的作用。需要进一步研究以调查它们在栀子中的作用,从而开发延缓花瓣衰老的工具。