Polavarapu Sailaja, Mani Arul M, Gundala Naveen K V, Hari Anasuya D, Bathina Siresha, Das Undurti N
Bio-Science Research Centre, Gayatri Vidya Parishad College of Engineering Campus, Madhurawada, Visakhapatnam-530048, Andhra Pradesh, India.
UND Life Sciences, 2020 S 360th St, # K-202, Federal Way, Washington, 98003, United States of America.
PLoS One. 2014 Dec 23;9(12):e114766. doi: 10.1371/journal.pone.0114766. eCollection 2014.
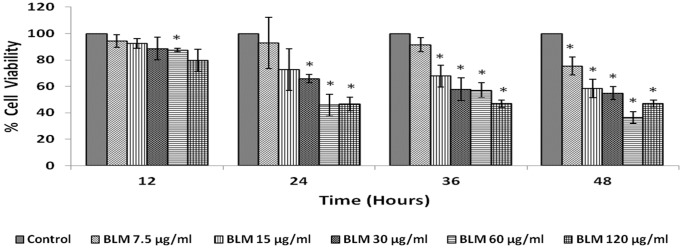
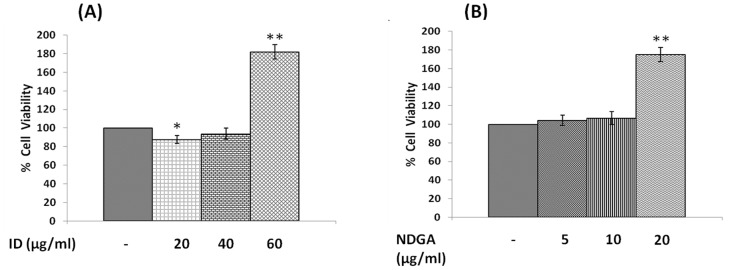
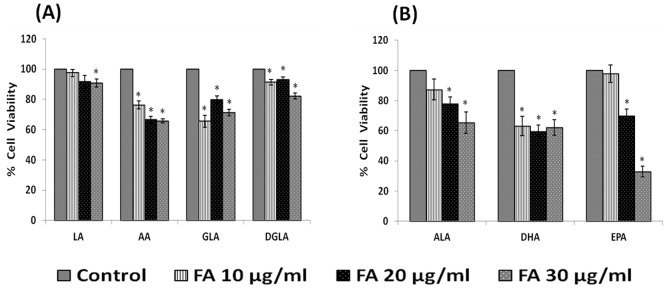
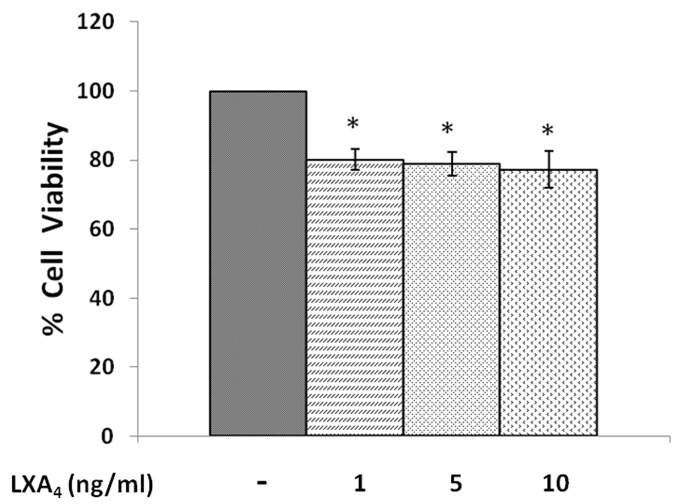
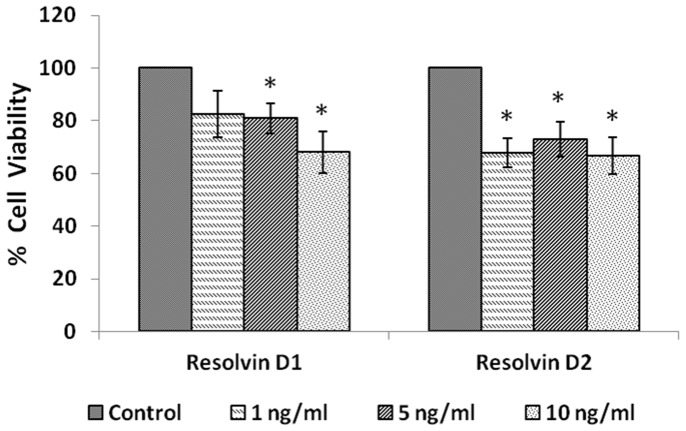
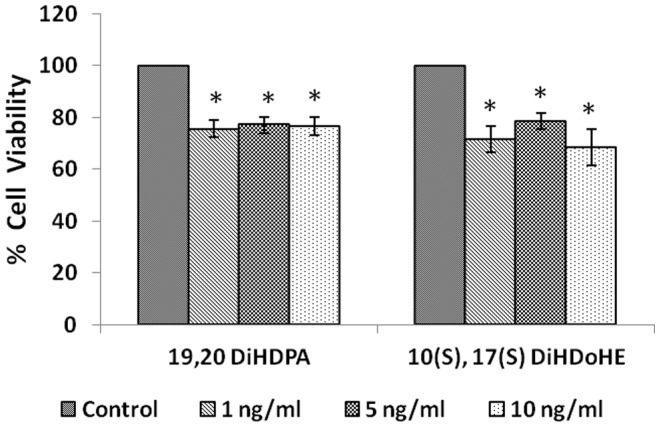
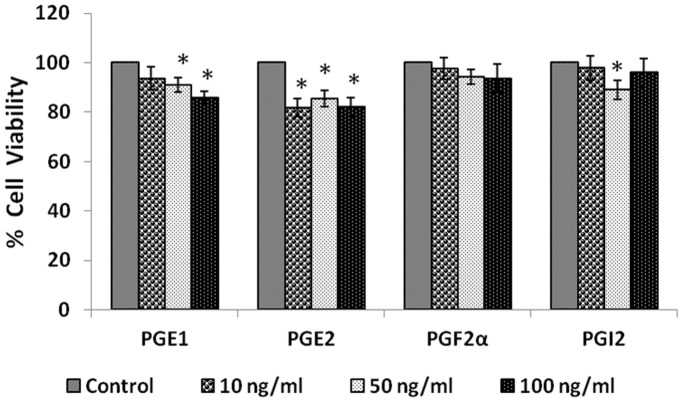
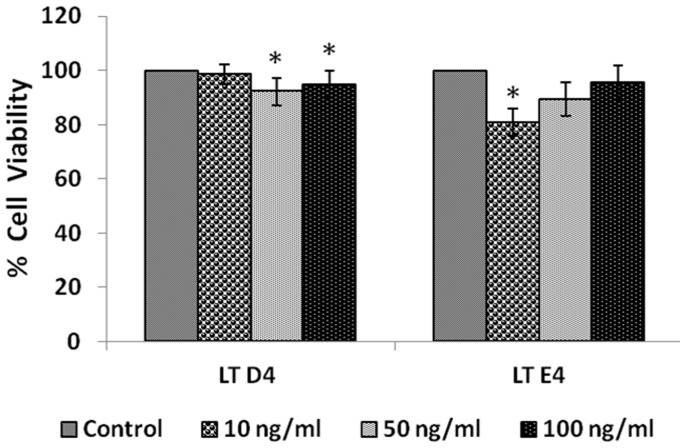
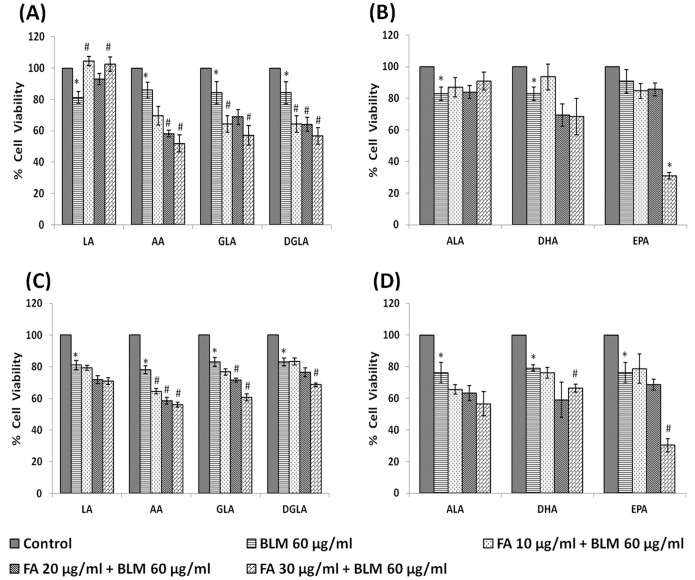
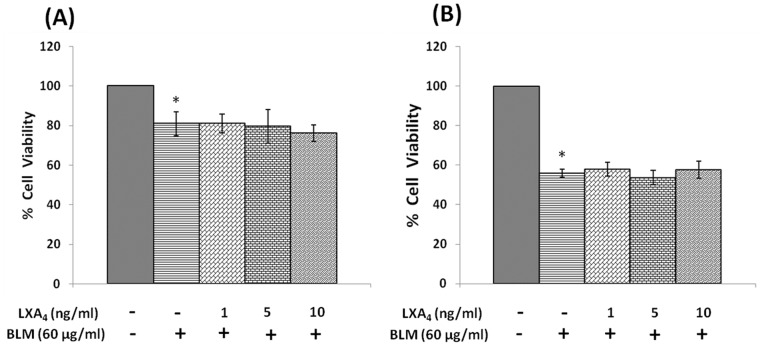
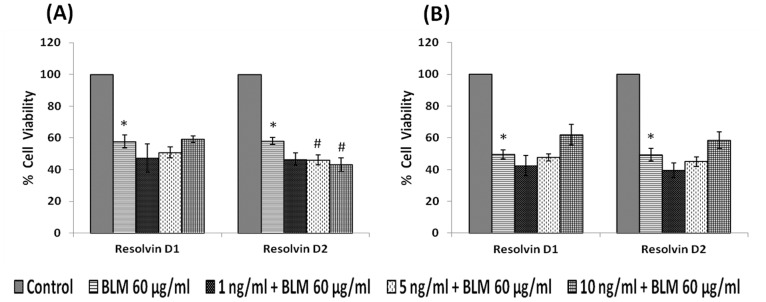
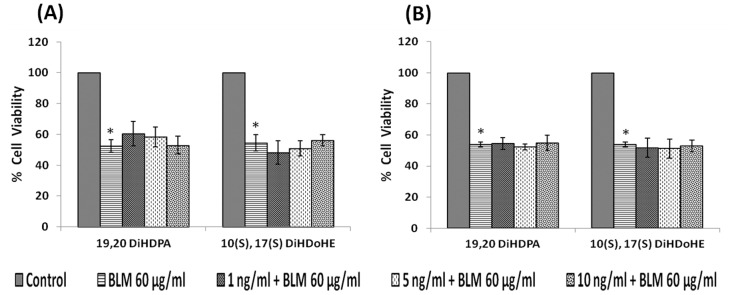
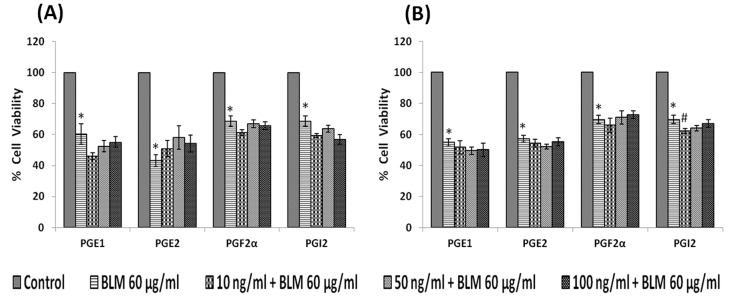
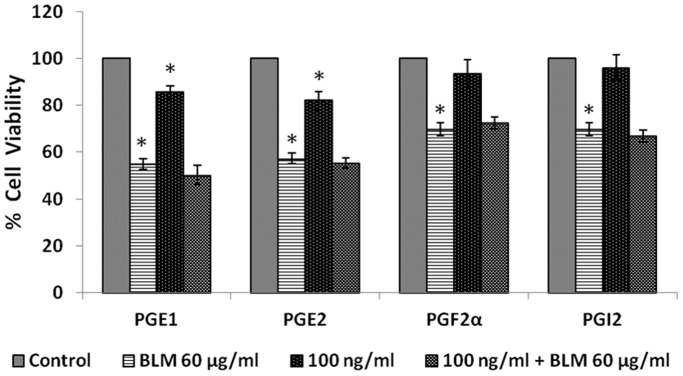
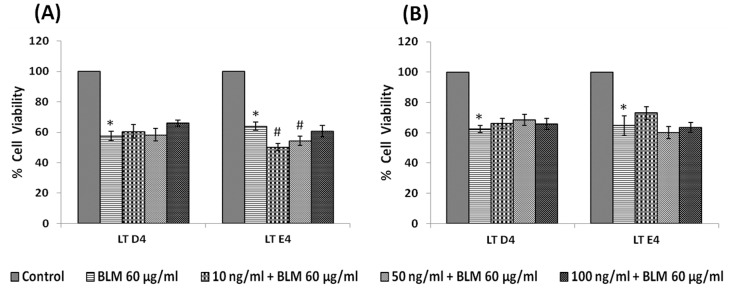
In the present study, we noted that bleomycin induced growth inhibitory action was augmented by all the polyunsaturated fatty acids (PUFAs) tested on human neuroblastoma IMR-32 (0.5 × 10(4) cells/100 µl of IMR) cells (EPA > DHA > ALA = GLA = AA > DGLA = LA: ∼ 60, 40, 30, 10-20% respectively) at the maximum doses used. Of all the prostaglandins (PGE1, PGE2, PGF2α, and PGI2) and leukotrienes (LTD4 and LTE4) tested; PGE1, PGE2 and LTD4 inhibited the growth of IMR-32 cells to a significant degree at the highest doses used. Lipoxin A4 (LXA4), 19,20-dihydroxydocosapentaenoate (19, 20 DiHDPA) and 10(S),17(S)-dihydroxy-4Z,7Z,11E,13Z,15E,19Z-docosahexaenoic acid (protectin: 10(S),17(S)DiHDoHE), metabolites of DHA, significantly inhibited the growth of IMR-32 cells. Pre-treatment with AA, GLA, DGLA and EPA and simultaneous treatment with all PUFAs used in the study augmented growth inhibitory action of bleomycin. Surprisingly, both indomethacin and nordihydroguaiaretic acid (NDGA) at 60 and 20 µg/ml respectively enhanced the growth of IMR-32 cells even in the presence of bleomycin. AA enhanced oxidant stress in IMR-32 cells as evidenced by an increase in lipid peroxides, superoxide dismutase levels and glutathione peroxidase activity. These results suggest that PUFAs suppress growth of human neuroblastoma cells, augment growth inhibitory action of bleomycin by enhancing formation of lipid peroxides and altering the status of anti-oxidants and, in all probability, increase the formation of lipoxins, resolvins and protectins from their respective precursors that possess growth inhibitory actions.
在本研究中,我们注意到,在所用的最大剂量下,博来霉素诱导的生长抑制作用在对人神经母细胞瘤IMR-32(0.5×10⁴细胞/100μl IMR)细胞测试的所有多不饱和脂肪酸(PUFA)作用下均增强(EPA>DHA>ALA = GLA = AA>DGLA = LA:分别约为60%、40%、30%、10%-20%)。在测试的所有前列腺素(PGE1、PGE2、PGF2α和PGI2)和白三烯(LTD4和LTE4)中,PGE1、PGE2和LTD4在所用的最高剂量下对IMR-32细胞的生长有显著抑制作用。DHA的代谢产物脂oxin A4(LXA4)、19,20-二羟基二十二碳五烯酸(19,20 DiHDPA)和10(S),17(S)-二羟基-4Z,7Z,11E,13Z,15E,19Z-二十二碳六烯酸(保护素:10(S),17(S)DiHDoHE)显著抑制IMR-32细胞的生长。用AA、GLA、DGLA和EPA预处理以及与本研究中使用的所有PUFA同时处理,增强了博来霉素的生长抑制作用。令人惊讶的是,即使在存在博来霉素的情况下,吲哚美辛和去甲二氢愈创木酸(NDGA)分别在60和20μg/ml时也增强了IMR-32细胞的生长。AA增加了IMR-32细胞中的氧化应激,脂质过氧化物、超氧化物歧化酶水平和谷胱甘肽过氧化物酶活性的增加证明了这一点。这些结果表明,PUFA抑制人神经母细胞瘤细胞的生长,通过增强脂质过氧化物的形成和改变抗氧化剂状态来增强博来霉素的生长抑制作用,并且很可能增加具有生长抑制作用的脂oxin、消退素和保护素从其各自前体的形成。