Rinerson Charles I, Rabara Roel C, Tripathi Prateek, Shen Qingxi J, Rushton Paul J
BMC Plant Biol. 2015 Feb 27;15:66. doi: 10.1186/s12870-015-0456-y.
The availability of increasing numbers of sequenced genomes has necessitated a re-evaluation of the evolution of the WRKY transcription factor family. Modern day plants descended from a charophyte green alga that colonized the land between 430 and 470 million years ago. The first charophyte genome sequence from Klebsormidium flaccidum filled a gap in the available genome sequences in the plant kingdom between unicellular green algae that typically have 1-3 WRKY genes and mosses that contain 30-40. WRKY genes have been previously found in non-plant species but their occurrence has been difficult to explain.
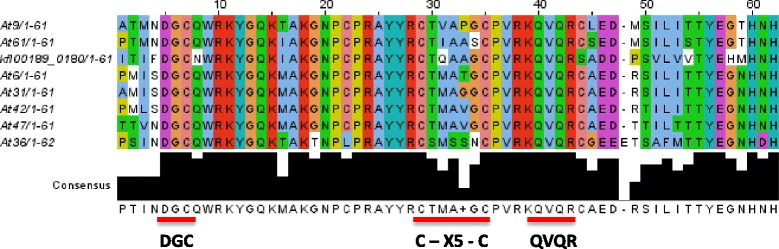
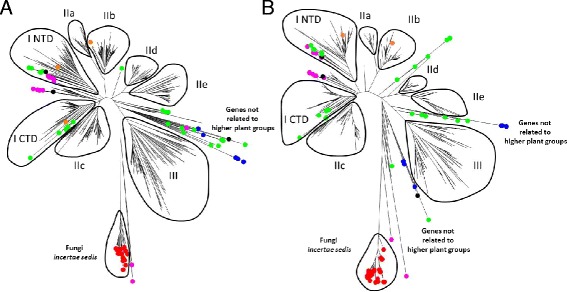
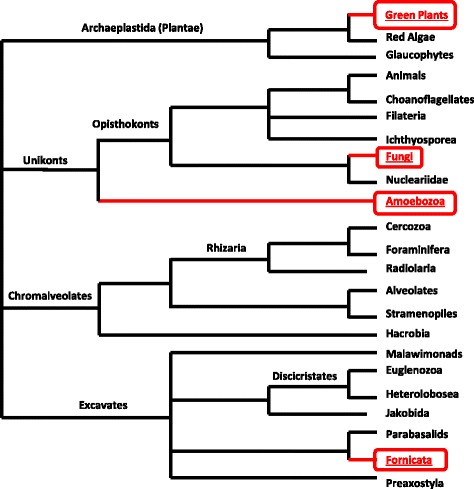
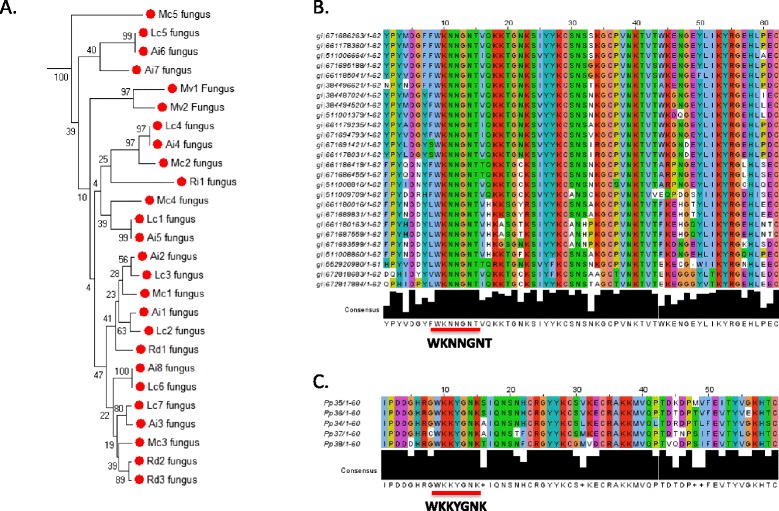
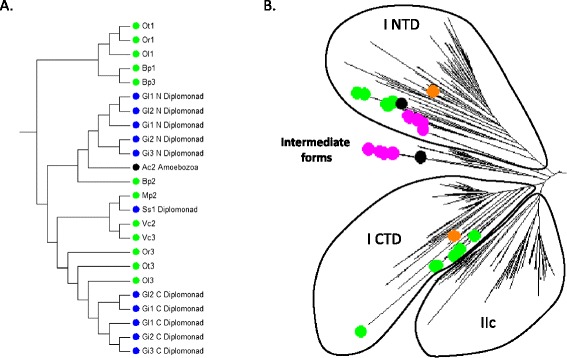
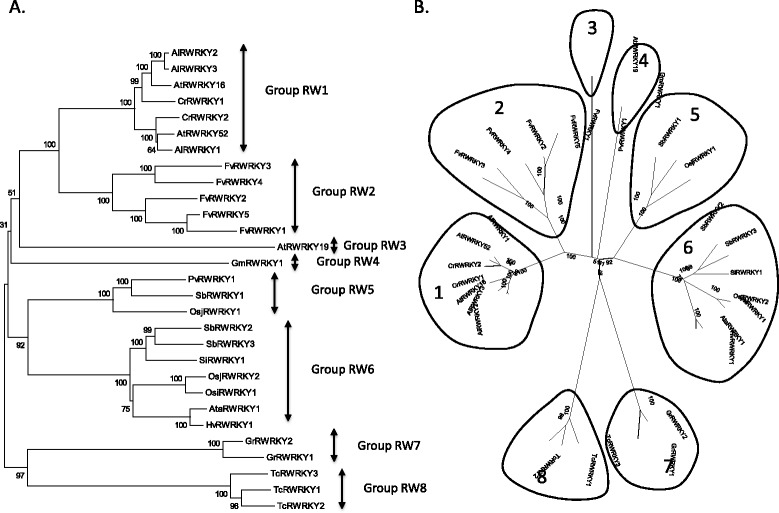
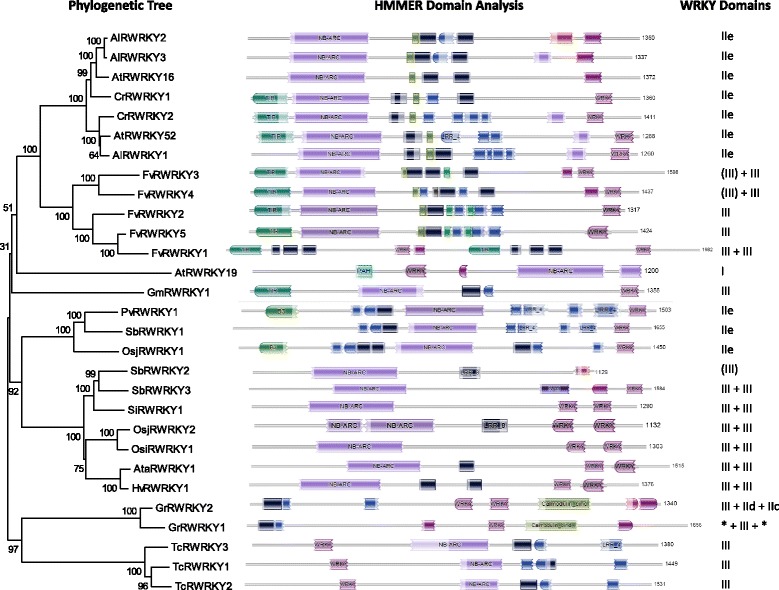
Only two WRKY genes are present in the Klebsormidium flaccidum genome and the presence of a Group IIb gene was unexpected because it had previously been thought that Group IIb WRKY genes first appeared in mosses. We found WRKY transcription factor genes outside of the plant lineage in some diplomonads, social amoebae, fungi incertae sedis, and amoebozoa. This patchy distribution suggests that lateral gene transfer is responsible. These lateral gene transfer events appear to pre-date the formation of the WRKY groups in flowering plants. Flowering plants contain proteins with domains typical for both resistance (R) proteins and WRKY transcription factors. R protein-WRKY genes have evolved numerous times in flowering plants, each type being restricted to specific flowering plant lineages. These chimeric proteins contain not only novel combinations of protein domains but also novel combinations and numbers of WRKY domains. Once formed, R protein WRKY genes may combine different components of signalling pathways that may either create new diversity in signalling or accelerate signalling by short circuiting signalling pathways.
We propose that the evolution of WRKY transcription factors includes early lateral gene transfers to non-plant organisms and the occurrence of algal WRKY genes that have no counterparts in flowering plants. We propose two alternative hypotheses of WRKY gene evolution: The "Group I Hypothesis" sees all WRKY genes evolving from Group I C-terminal WRKY domains. The alternative "IIa + b Separate Hypothesis" sees Groups IIa and IIb evolving directly from a single domain algal gene separate from the Group I-derived lineage.
越来越多已测序基因组的可得性使得对WRKY转录因子家族的进化进行重新评估成为必要。现代植物起源于一种轮藻绿藻,这种绿藻在4.3亿至4.7亿年前殖民了陆地。来自柔弱刚毛藻的首个轮藻基因组序列填补了植物界可用基因组序列中的一个空白,该空白存在于通常具有1 - 3个WRKY基因的单细胞绿藻和含有30 - 40个WRKY基因的苔藓之间。WRKY基因此前已在非植物物种中被发现,但其出现难以解释。
柔弱刚毛藻基因组中仅存在两个WRKY基因,且IIb组基因的存在出乎意料,因为此前一直认为IIb组WRKY基因首次出现在苔藓中。我们在一些双滴虫、社会性变形虫、未定类真菌和变形虫门中发现了植物谱系之外的WRKY转录因子基因。这种零散的分布表明是横向基因转移导致的。这些横向基因转移事件似乎早于开花植物中WRKY组的形成。开花植物含有兼具抗性(R)蛋白和WRKY转录因子典型结构域的蛋白质。R蛋白 - WRKY基因在开花植物中已多次进化,每种类型都局限于特定的开花植物谱系。这些嵌合蛋白不仅包含蛋白质结构域的新组合,还包含WRKY结构域的新组合和数量。一旦形成,R蛋白WRKY基因可能会组合信号通路的不同组成部分,这可能会在信号传导中创造新的多样性或通过使信号通路短路来加速信号传导。
我们提出WRKY转录因子的进化包括早期向非植物生物的横向基因转移以及开花植物中不存在对应物的藻类WRKY基因的出现。我们提出了WRKY基因进化的两种替代假说:“I组假说”认为所有WRKY基因都从I组C末端WRKY结构域进化而来。另一种“IIa + b 独立假说”认为IIa组和IIb组直接从与I组衍生谱系分开的单个结构域藻类基因进化而来。