Van Steenberge Maarten, Pariselle Antoine, Huyse Tine, Volckaert Filip A M, Snoeks Jos, Vanhove Maarten P M
Biology Department, Royal Museum for Central Africa, Tervuren, Belgium; Laboratory of Biodiversity and Evolutionary Genomics, Department of Biology, University of Leuven, Leuven, Belgium; Institute of Zoology, University of Graz, Graz, Austria.
Institut des Sciences de l'Évolution, IRD-CNRS-Université Montpellier, Montpellier, France.
PLoS One. 2015 Apr 29;10(4):e0124474. doi: 10.1371/journal.pone.0124474. eCollection 2015.
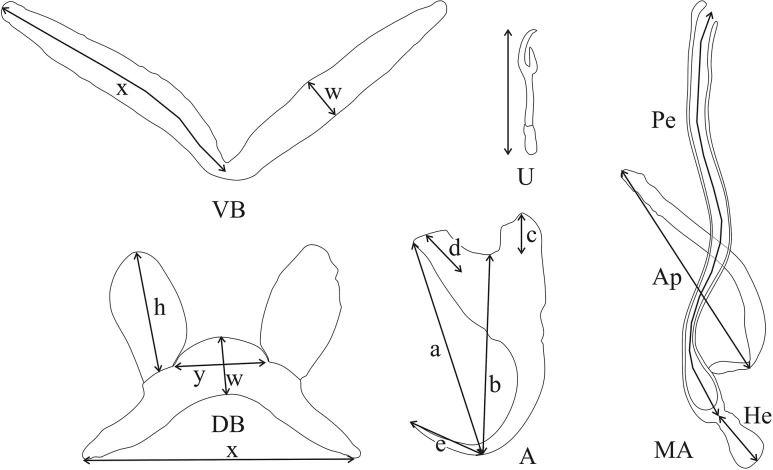
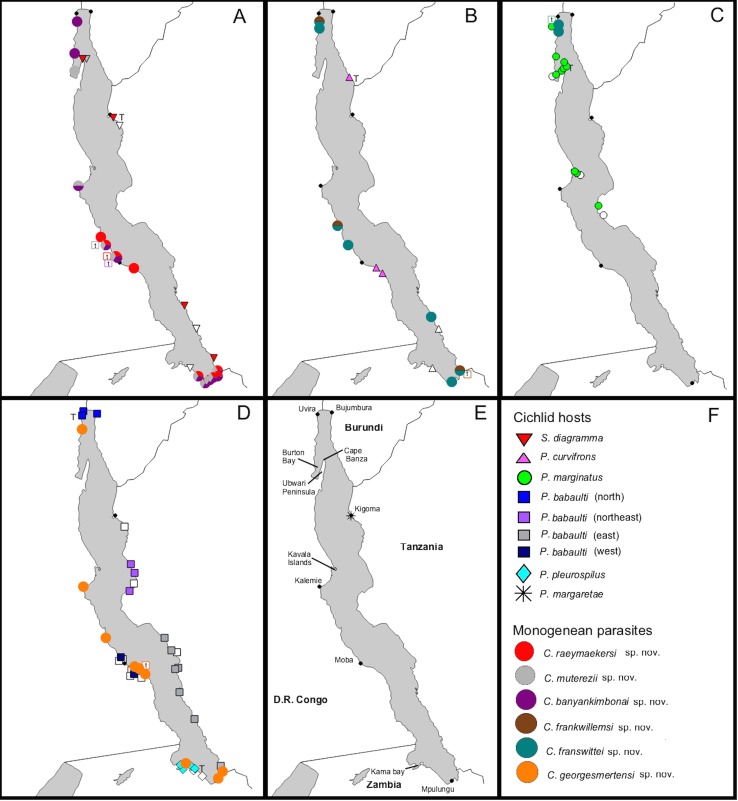
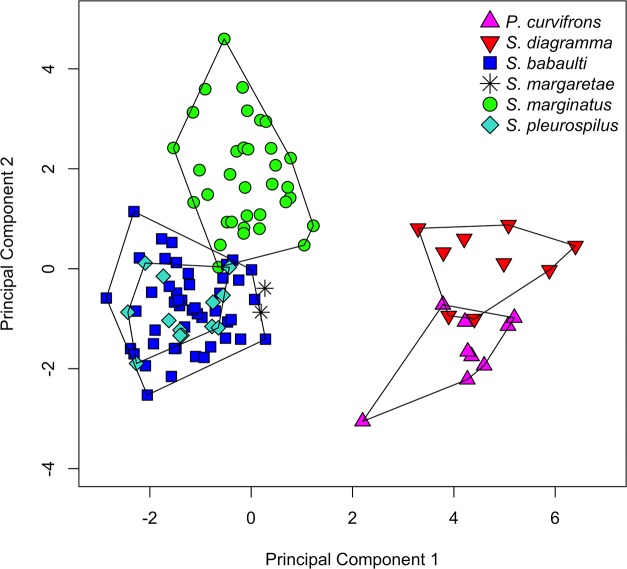
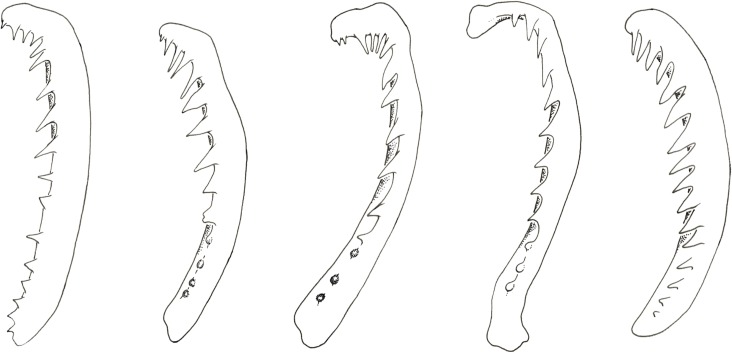
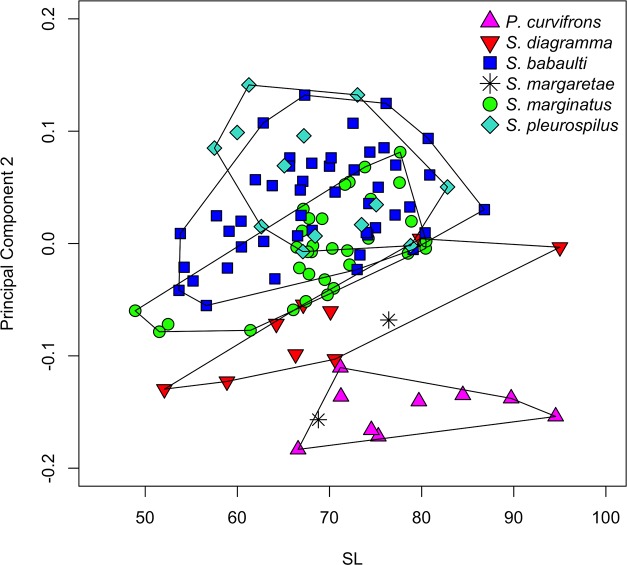
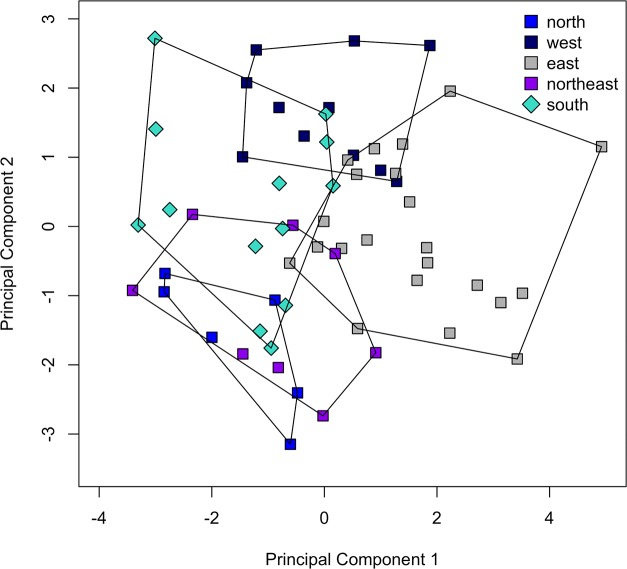
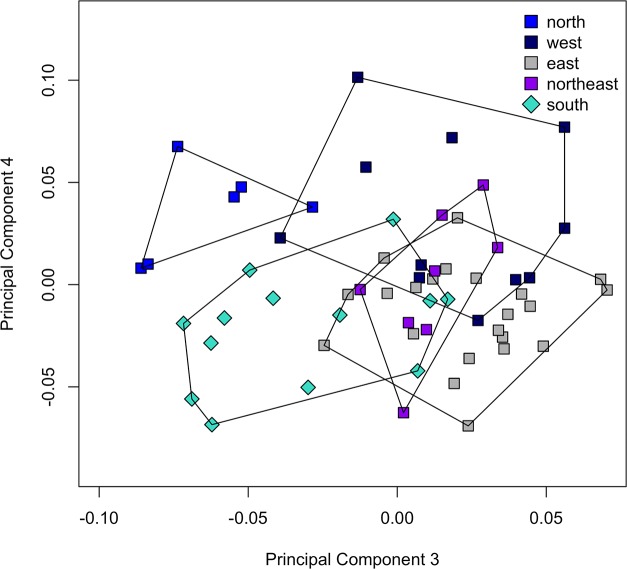
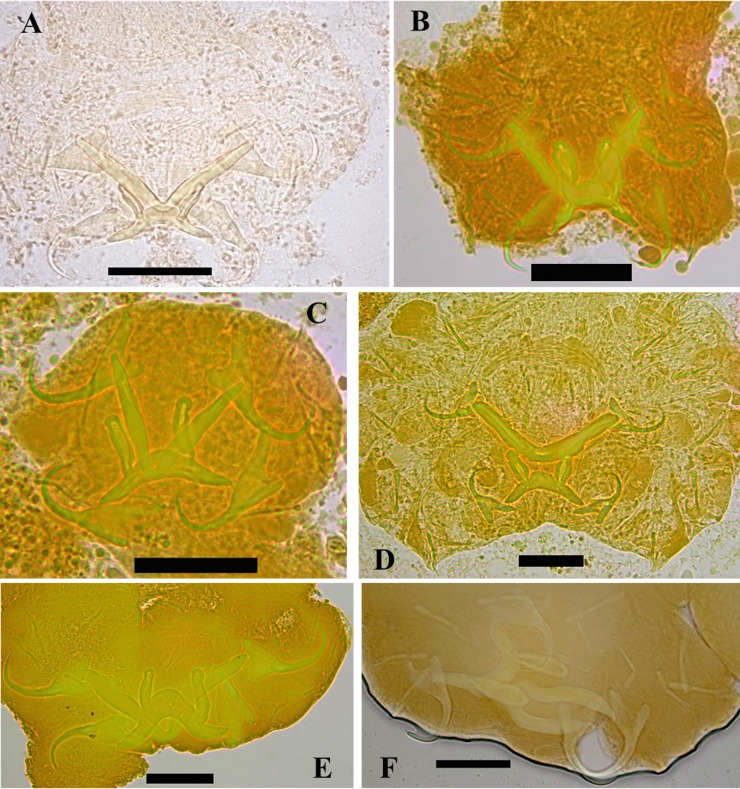
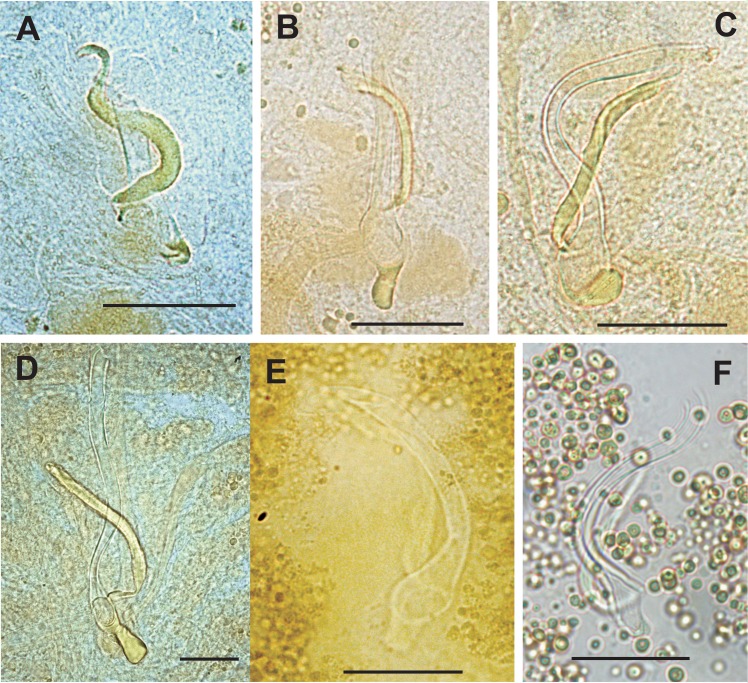
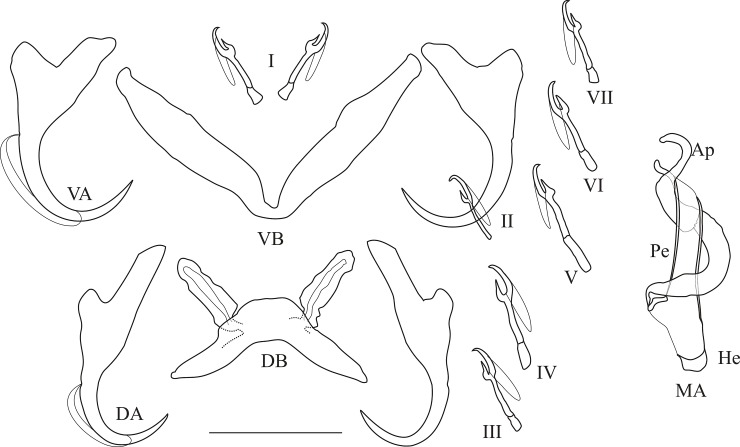
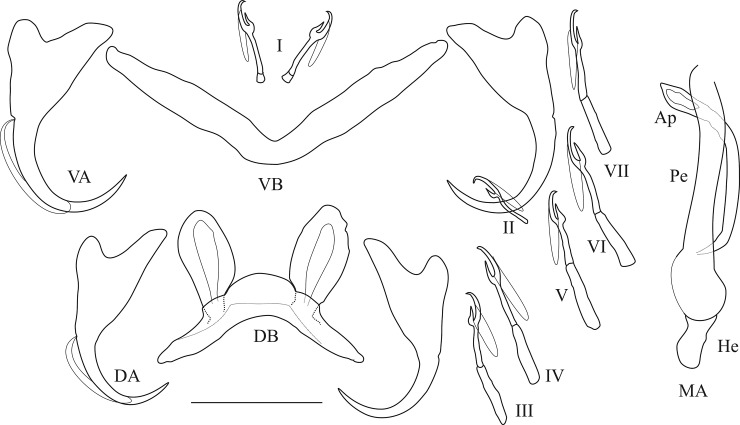
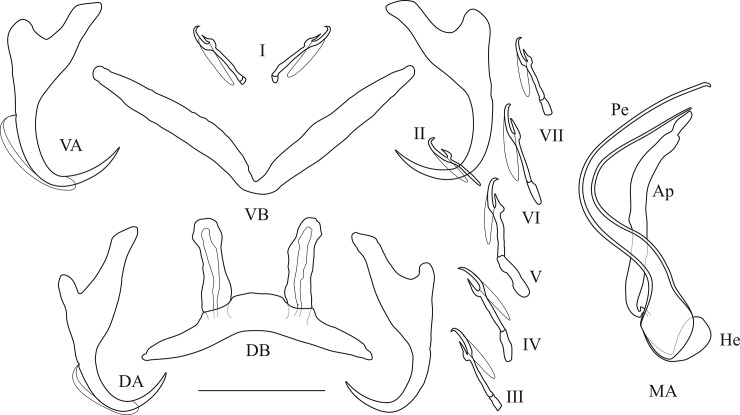
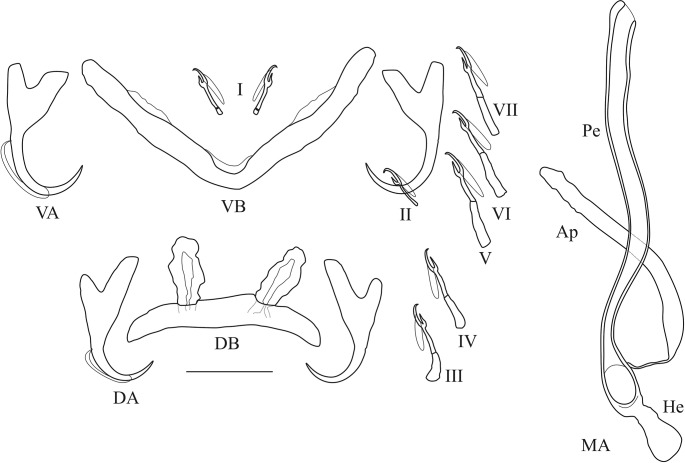
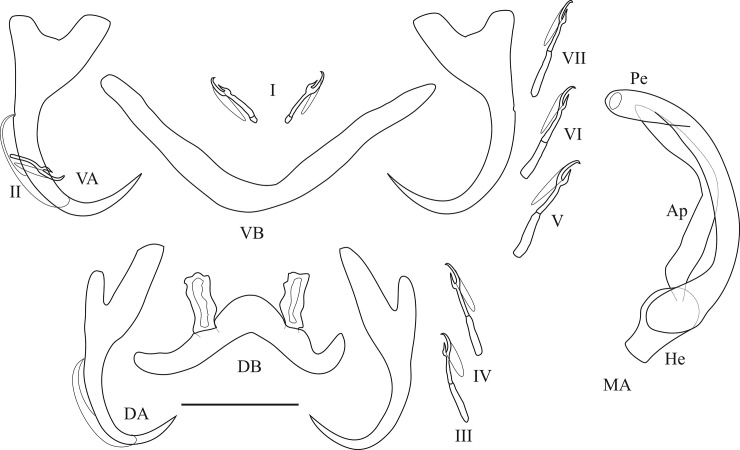
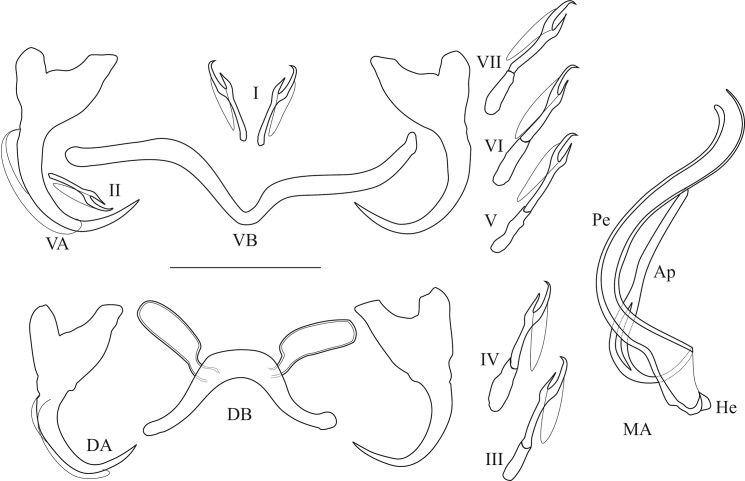
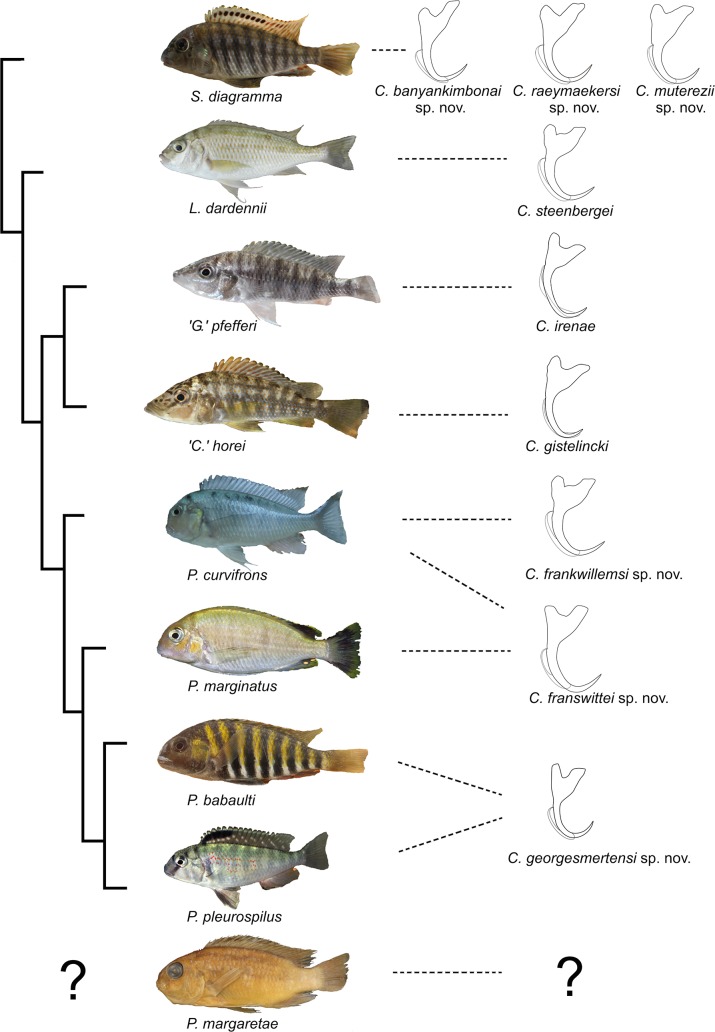
The unparalleled biodiversity of Lake Tanganyika (Africa) has fascinated biologists for over a century; its unique cichlid communities are a preferred model for evolutionary research. Although species delineation is, in most cases, relatively straightforward, higher-order classifications were shown not to agree with monophyletic groups. Here, traditional morphological methods meet their limitations. A typical example are the tropheine cichlids currently belonging to Simochromis and Pseudosimochromis. The affiliations of these widespread and abundant cichlids are poorly understood. Molecular work suggested that genus and species boundaries should be revised. Moreover, previous morphological results indicated that intraspecific variation should be considered to delineate species in Lake Tanganyika cichlids. We review the genera Simochromis and Pseudosimochromis using an integrative approach. Besides a morphometric study and a barcoding approach, monogenean Cichlidogyrus (Platyhelminthes: Ancyrocephalidae) gill parasites, often highly species-specific, are used as complementary markers. Six new species are described. Cichlidogyrus raeymaekersi sp. nov., C. muterezii sp. nov. and C. banyankimbonai sp. nov. infect S. diagramma. Cichlidogyrus georgesmertensi sp. nov. was found on S. babaulti and S. pleurospilus, C. franswittei sp. nov. on both S. marginatus and P. curvifrons and C. frankwillemsi sp. nov. only on P. curvifrons. As relatedness between Cichlidogyrus species usually reflects relatedness between hosts, we considered Simochromis monotypic because the three Cichlidogyrus species found on S. diagramma belonged to a different morphotype than those found on the other Simochromis. The transfer of S. babaulti, S. marginatus, S. pleurospilus and S. margaretae to Pseudosimochromis was justified by the similarity of their Cichlidogyrus fauna and the intermediate morphology of S. margaretae. Finally parasite data also supported the synonymy between S. pleurospilus and S. babaulti, a species that contains a large amount of geographical morphological variation.
非洲坦噶尼喀湖无与伦比的生物多样性已吸引生物学家长达一个多世纪;其独特的丽鱼科群落是进化研究的首选模式。尽管在大多数情况下,物种划分相对简单,但高阶分类结果显示与单系群并不一致。在此,传统形态学方法遇到了其局限性。一个典型例子是目前属于似丽鱼属(Simochromis)和伪似丽鱼属(Pseudosimochromis)的曲唇丽鱼科丽鱼。这些分布广泛且数量众多的丽鱼的分类归属尚不清楚。分子研究表明,属和种的界限应予以修订。此外,先前的形态学研究结果表明,在划分坦噶尼喀湖丽鱼科物种时应考虑种内变异。我们采用综合方法对似丽鱼属和伪似丽鱼属进行了综述。除了形态测量研究和条形码方法外,单殖吸虫丽鱼三代虫(扁形动物门:锚首虫科)鳃寄生虫(通常具有高度物种特异性)被用作补充标记。描述了六个新物种。雷马克尔斯丽鱼三代虫(Cichlidogyrus raeymaekersi sp. nov.)、穆特雷齐丽鱼三代虫(C. muterezii sp. nov.)和班扬金博奈丽鱼三代虫(C. banyankimbonai sp. nov.)感染图氏似丽鱼(S. diagramma)。乔治斯默滕斯丽鱼三代虫(Cichlidogyrus georgesmertensi sp. nov.)在巴氏似丽鱼(S. babaulti)和肋斑似丽鱼(S. pleurospilus)上被发现,弗兰斯维特丽鱼三代虫(C. franswittei sp. nov.)在缘边似丽鱼(S. marginatus)和弯额伪似丽鱼(P. curvifrons)上均有发现,而弗兰克维勒姆斯丽鱼三代虫(C. frankwillemsi sp. nov.)仅在弯额伪似丽鱼上被发现。由于丽鱼三代虫物种之间的亲缘关系通常反映宿主之间的亲缘关系,我们认为似丽鱼属为单型属,因为在图氏似丽鱼上发现的三种丽鱼三代虫与在其他似丽鱼属鱼类上发现的属于不同形态类型。将巴氏似丽鱼、缘边似丽鱼、肋斑似丽鱼和玛格丽塔似丽鱼转移到伪似丽鱼属是合理的,这是基于它们的丽鱼三代虫动物群的相似性以及玛格丽塔似丽鱼的中间形态。最后,寄生虫数据也支持肋斑似丽鱼和巴氏似丽鱼同义,巴氏似丽鱼包含大量地理形态变异。