Talavera David, Lovell Simon C, Whelan Simon
Faculty of Life Sciences, University of Manchester, Manchester, United Kingdom
Faculty of Life Sciences, University of Manchester, Manchester, United Kingdom.
Mol Biol Evol. 2015 Sep;32(9):2456-68. doi: 10.1093/molbev/msv109. Epub 2015 May 4.
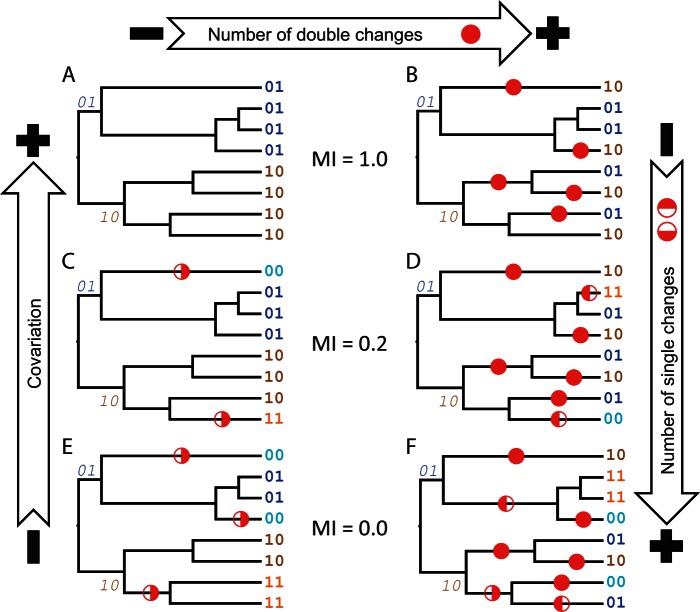
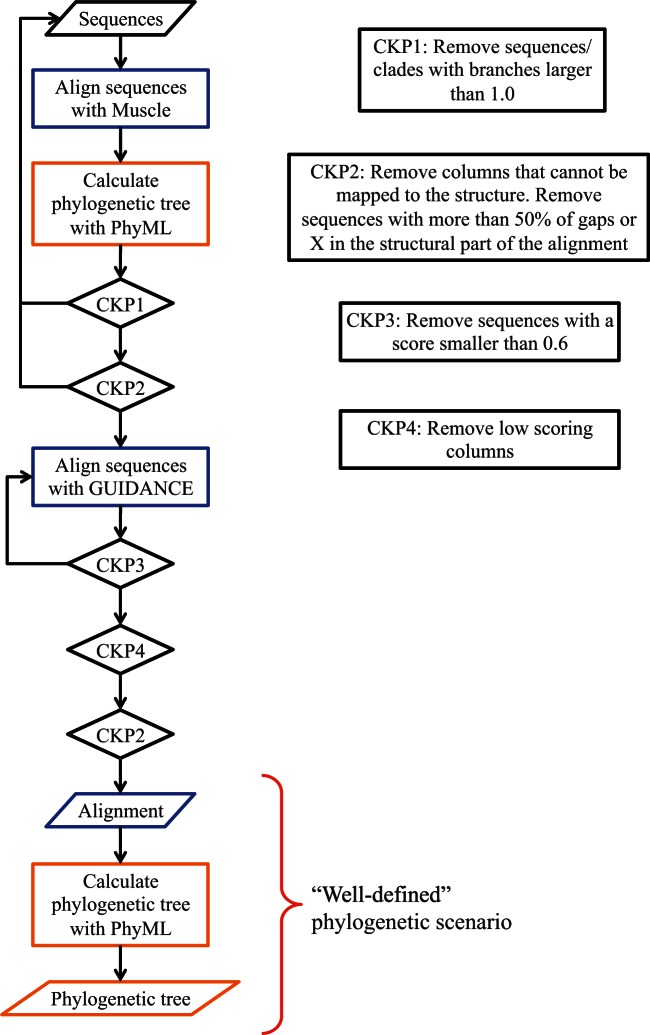
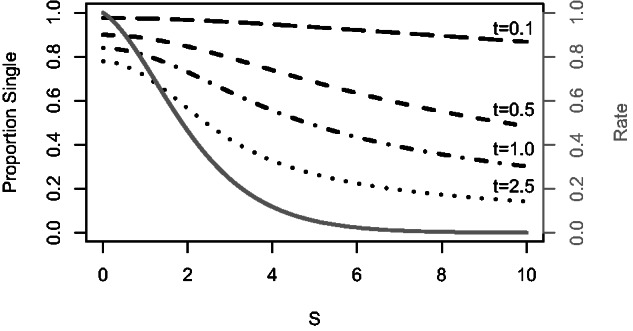
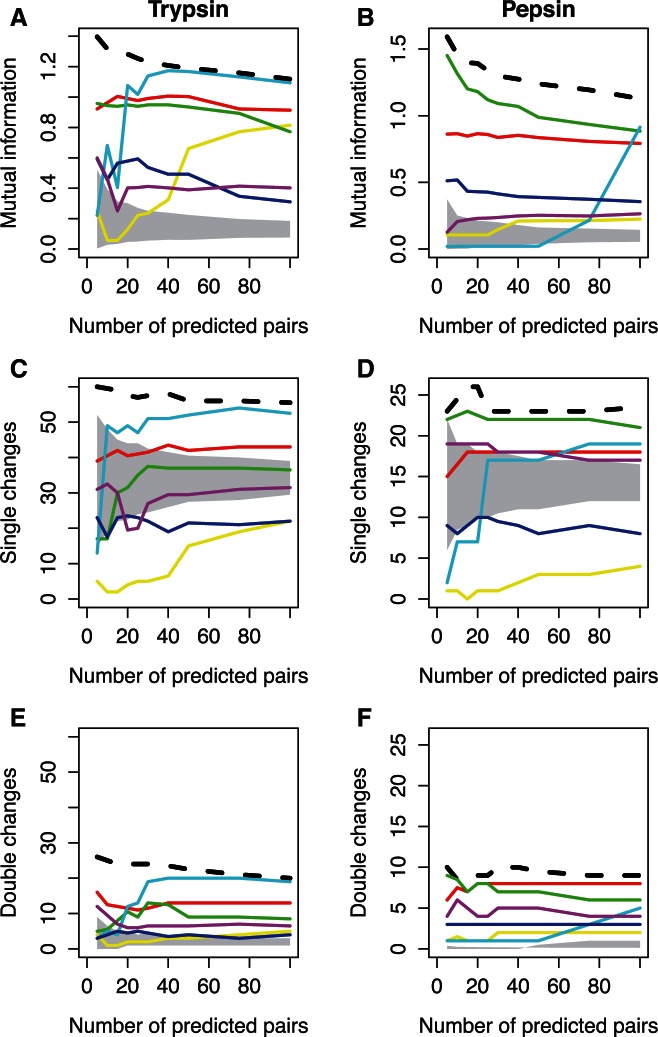
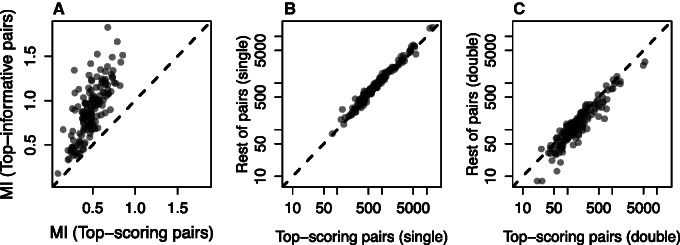
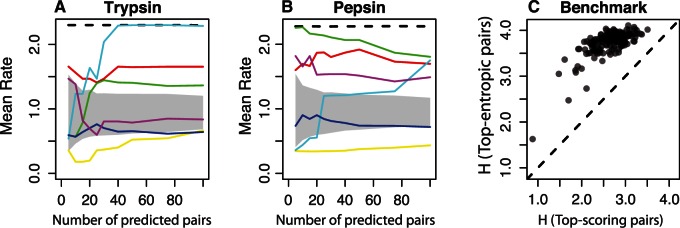
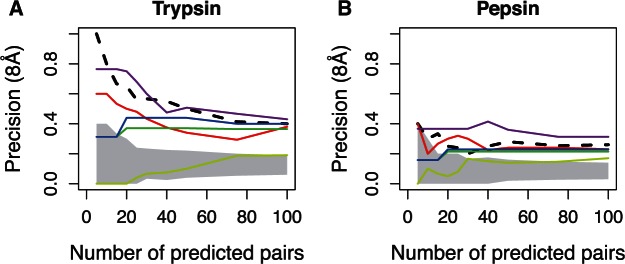
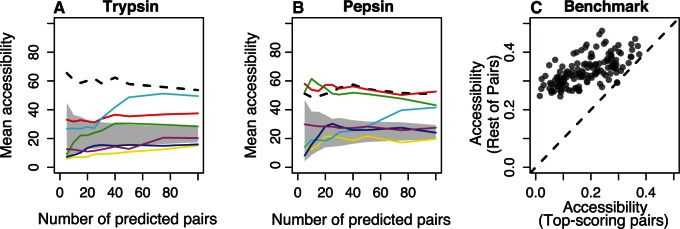
Recent developments in the analysis of amino acid covariation are leading to breakthroughs in protein structure prediction, protein design, and prediction of the interactome. It is assumed that observed patterns of covariation are caused by molecular coevolution, where substitutions at one site affect the evolutionary forces acting at neighboring sites. Our theoretical and empirical results cast doubt on this assumption. We demonstrate that the strongest coevolutionary signal is a decrease in evolutionary rate and that unfeasibly long times are required to produce coordinated substitutions. We find that covarying substitutions are mostly found on different branches of the phylogenetic tree, indicating that they are independent events that may or may not be attributable to coevolution. These observations undermine the hypothesis that molecular coevolution is the primary cause of the covariation signal. In contrast, we find that the pairs of residues with the strongest covariation signal tend to have low evolutionary rates, and that it is this low rate that gives rise to the covariation signal. Slowly evolving residue pairs are disproportionately located in the protein's core, which explains covariation methods' ability to detect pairs of residues that are close in three dimensions. These observations lead us to propose the "coevolution paradox": The strength of coevolution required to cause coordinated changes means the evolutionary rate is so low that such changes are highly unlikely to occur. As modern covariation methods may lead to breakthroughs in structural genomics, it is critical to recognize their biases and limitations.
氨基酸共变分析的最新进展正在推动蛋白质结构预测、蛋白质设计和相互作用组预测方面取得突破。人们认为观察到的共变模式是由分子协同进化引起的,其中一个位点的替换会影响作用于相邻位点的进化力量。我们的理论和实证结果对这一假设提出了质疑。我们证明,最强的协同进化信号是进化速率的降低,并且产生协同替换需要长得不可行的时间。我们发现共变替换大多出现在系统发育树的不同分支上,这表明它们是独立事件,可能归因于协同进化,也可能不归因于协同进化。这些观察结果削弱了分子协同进化是共变信号主要原因的假设。相比之下,我们发现具有最强共变信号的残基对往往进化速率较低,正是这种低速率产生了共变信号。缓慢进化的残基对不成比例地位于蛋白质的核心区域,这解释了共变方法检测三维空间中靠近的残基对的能力。这些观察结果促使我们提出“协同进化悖论”:导致协同变化所需的协同进化强度意味着进化速率极低,以至于这种变化极不可能发生。由于现代共变方法可能会在结构基因组学方面取得突破,认识到它们的偏差和局限性至关重要。