Rogala Kacper B, Dynes Nicola J, Hatzopoulos Georgios N, Yan Jun, Pong Sheng Kai, Robinson Carol V, Deane Charlotte M, Gönczy Pierre, Vakonakis Ioannis
Department of Biochemistry, University of Oxford, Oxford, United Kingdom.
Swiss Institute for Experimental Cancer Research, School of Life Sciences, Swiss Federal Institute of Technology, Lausanne, Switzerland.
Elife. 2015 May 29;4:e07410. doi: 10.7554/eLife.07410.
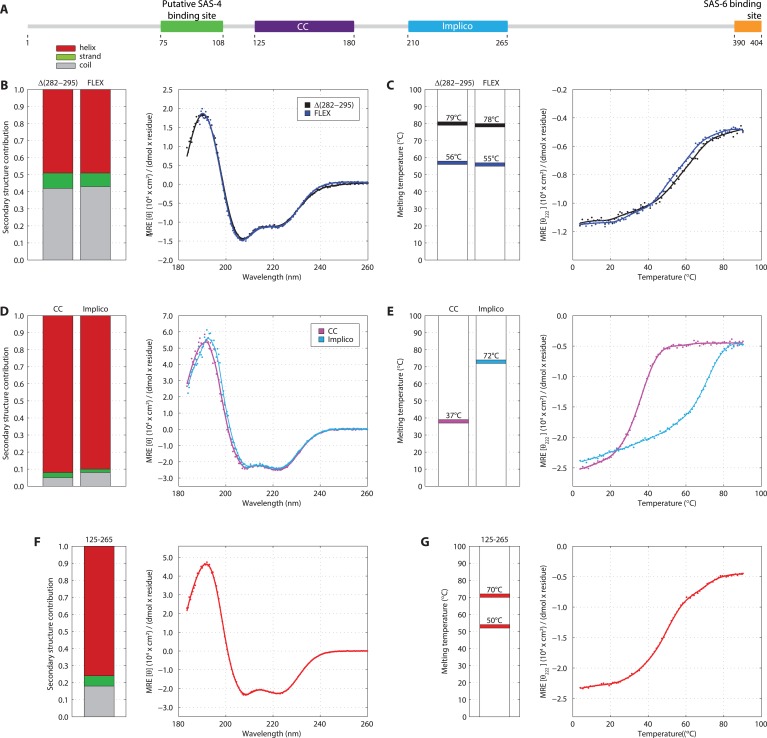
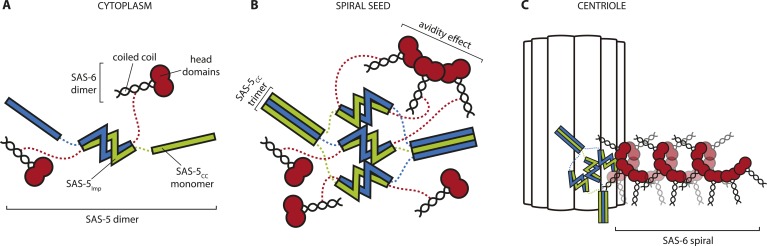
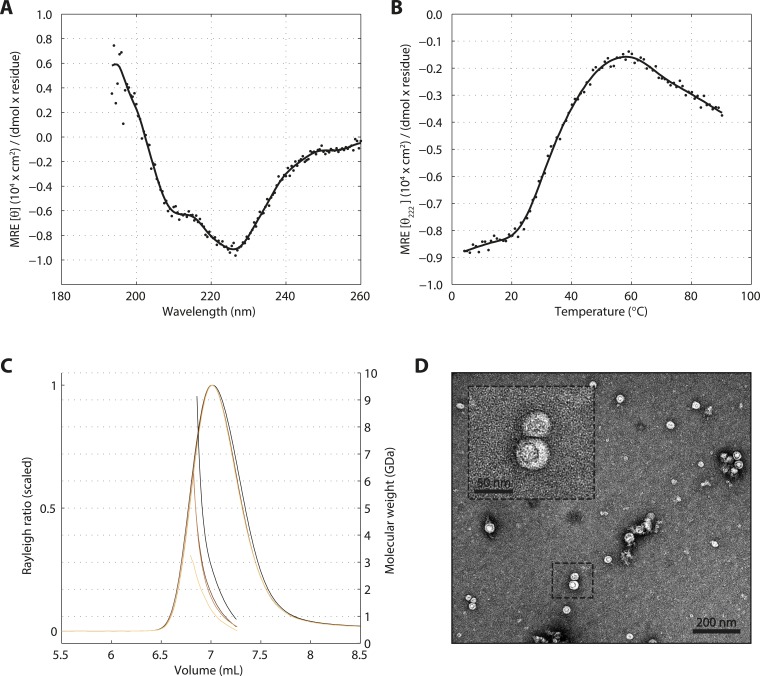
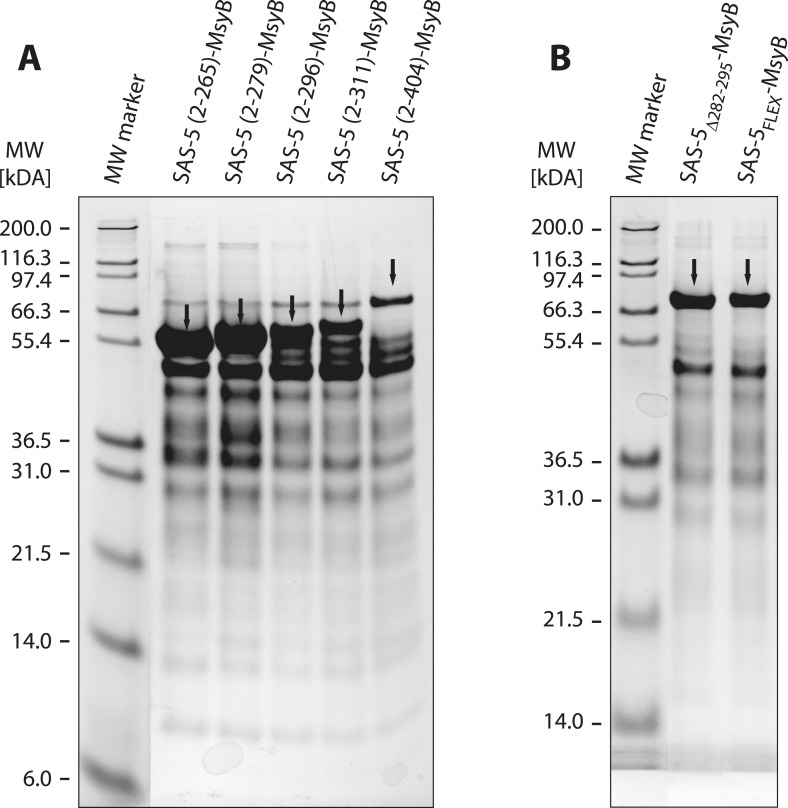
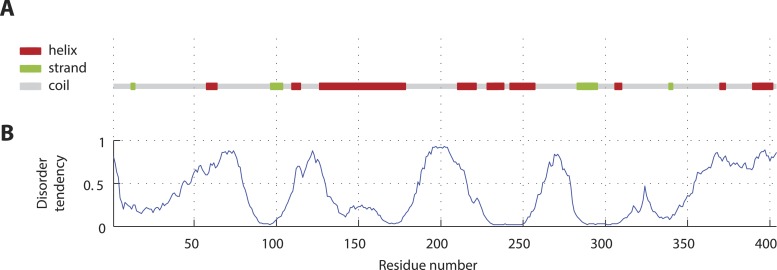
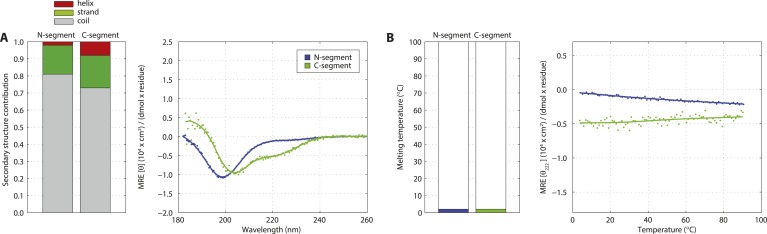
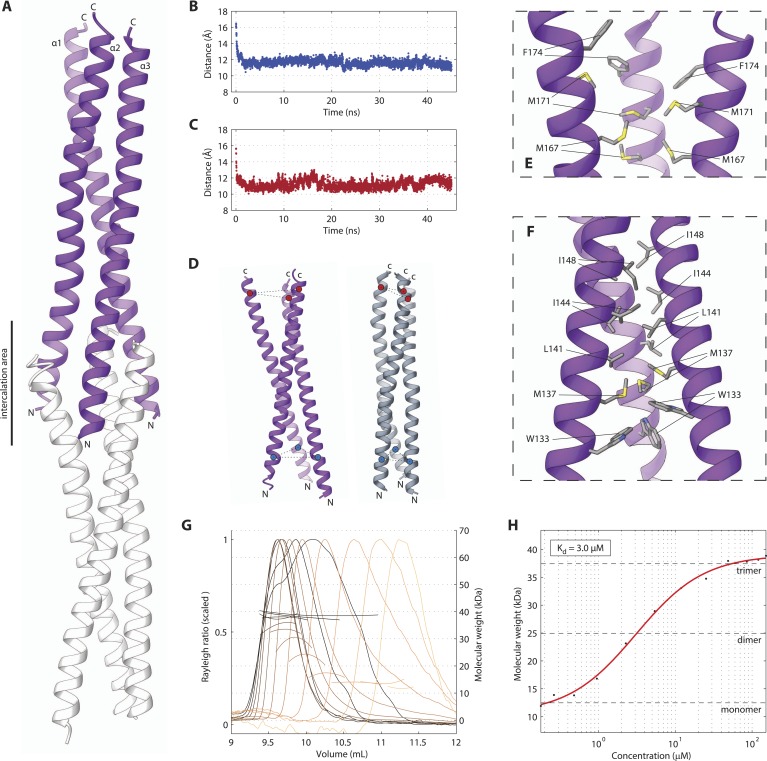
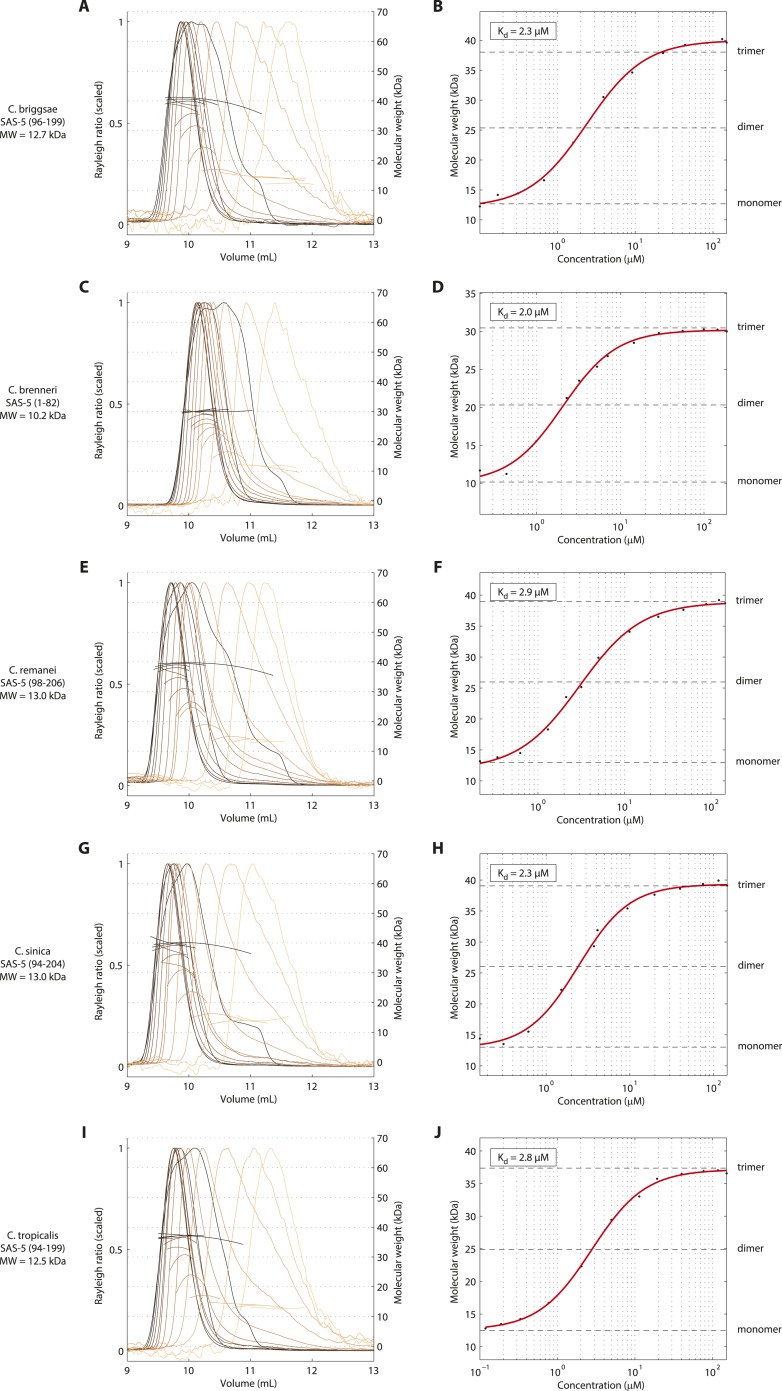
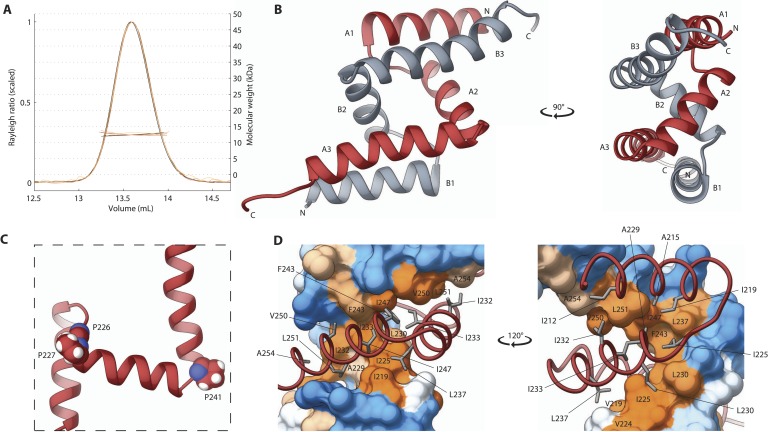
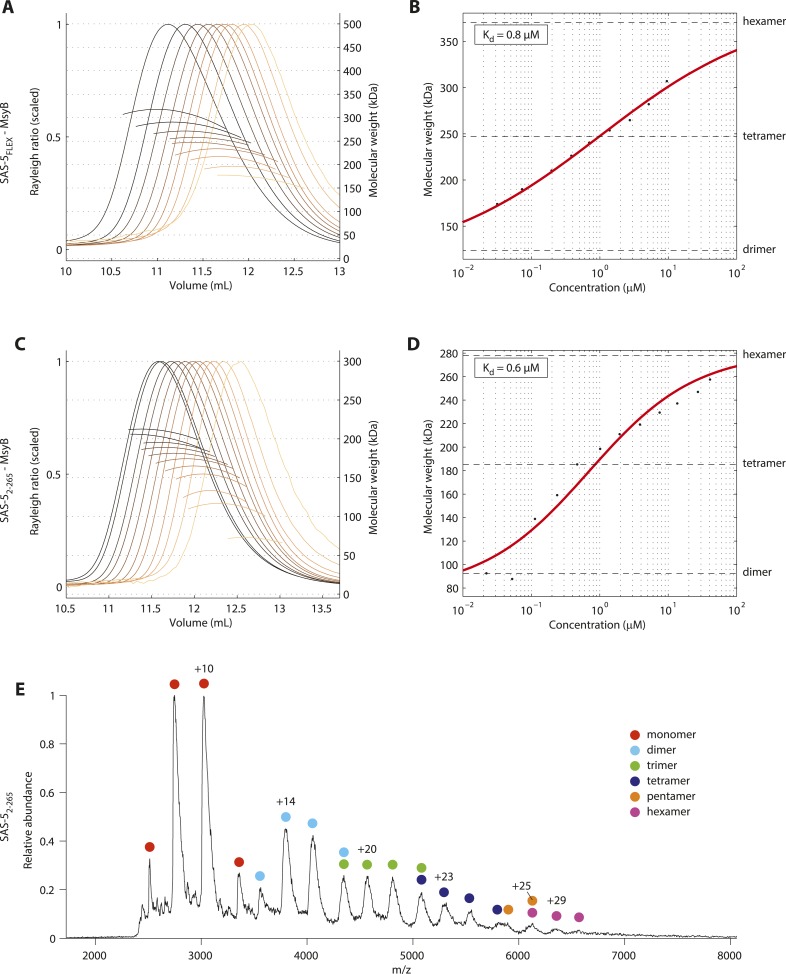
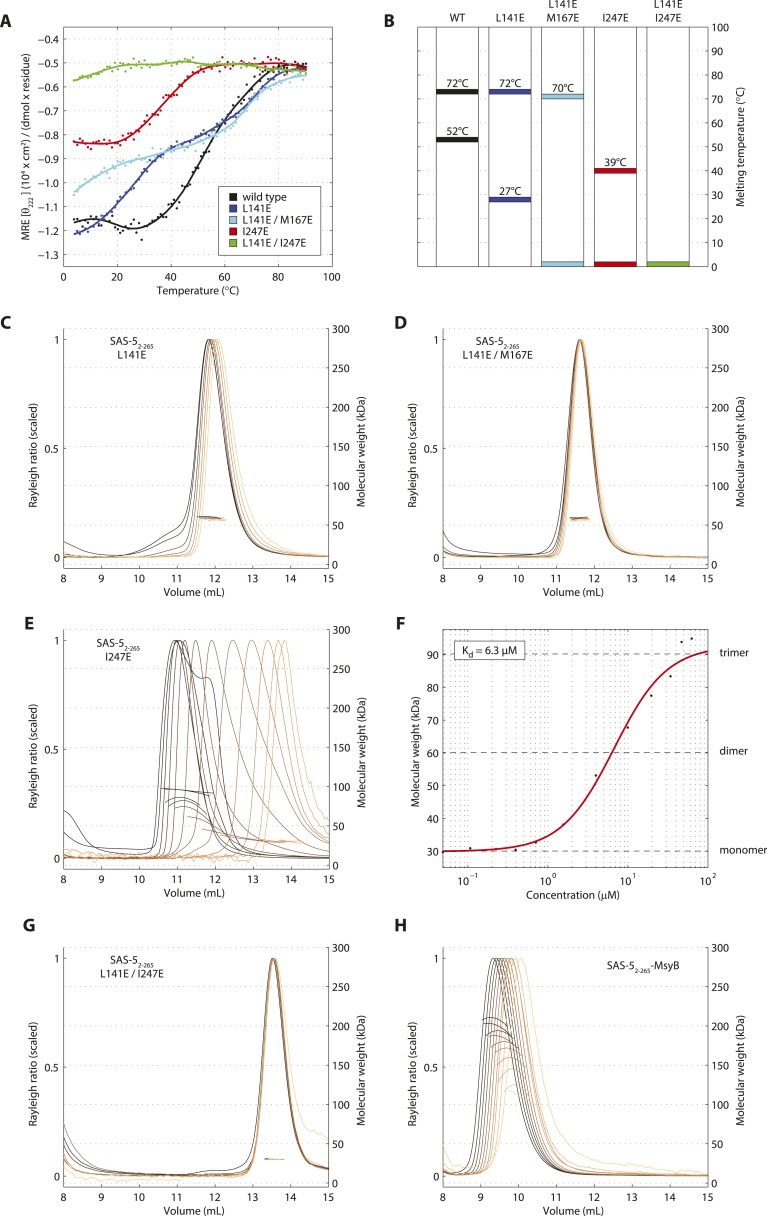
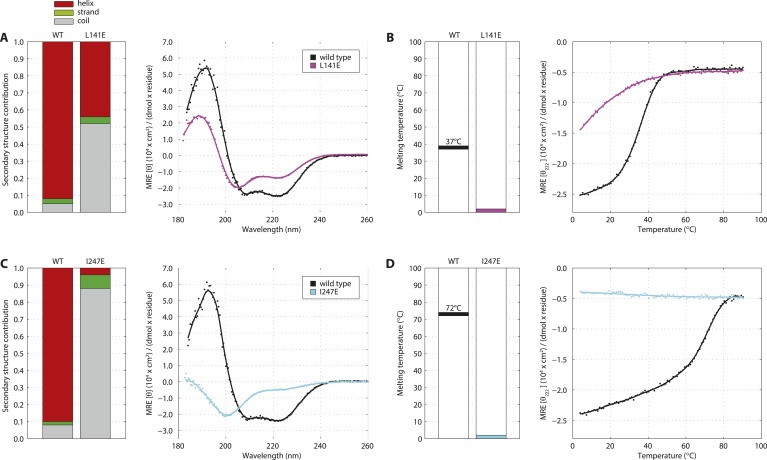
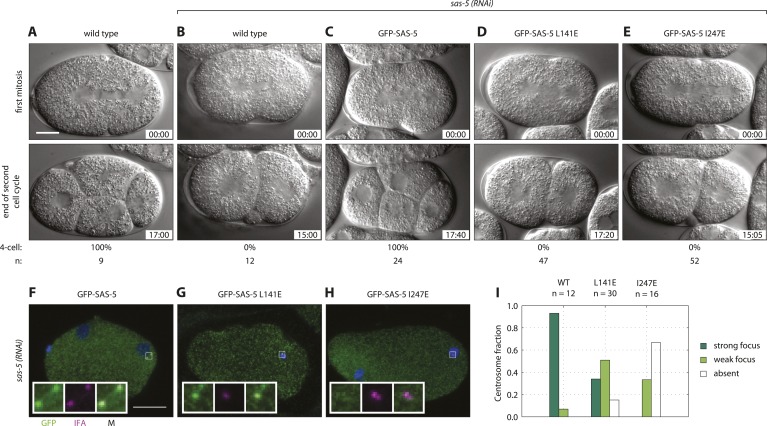
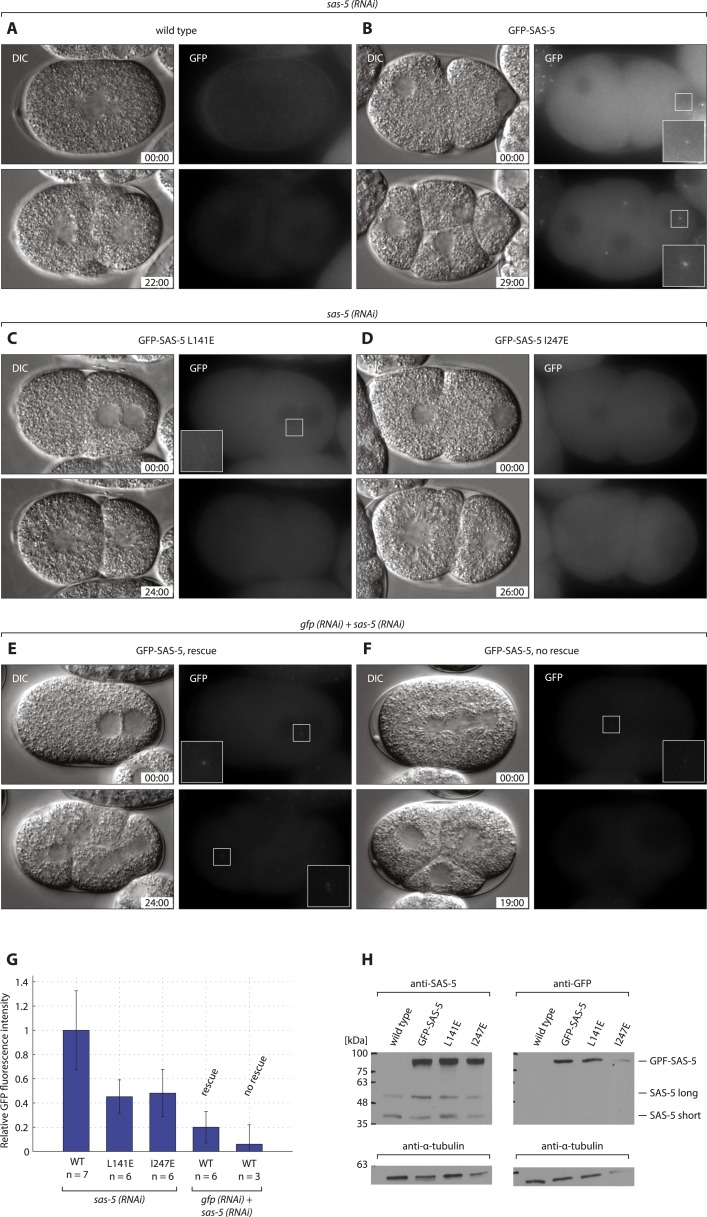
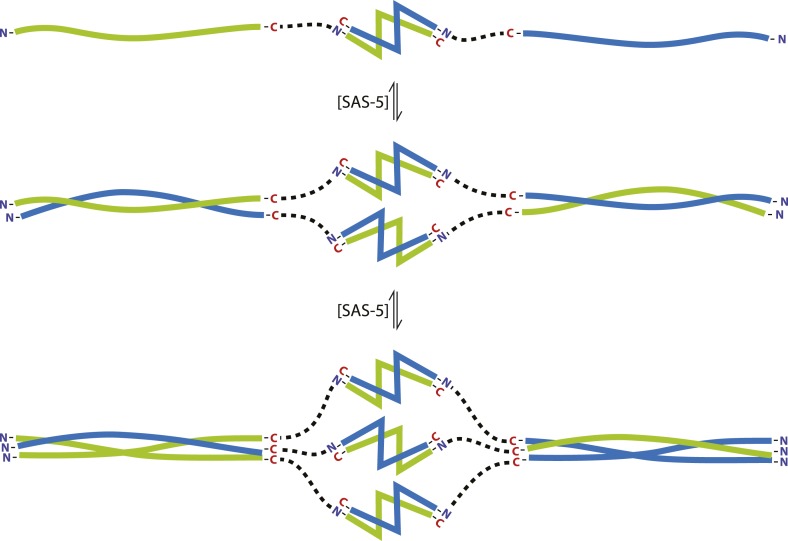
Centrioles are microtubule-based organelles crucial for cell division, sensing and motility. In Caenorhabditis elegans, the onset of centriole formation requires notably the proteins SAS-5 and SAS-6, which have functional equivalents across eukaryotic evolution. Whereas the molecular architecture of SAS-6 and its role in initiating centriole formation are well understood, the mechanisms by which SAS-5 and its relatives function is unclear. Here, we combine biophysical and structural analysis to uncover the architecture of SAS-5 and examine its functional implications in vivo. Our work reveals that two distinct self-associating domains are necessary to form higher-order oligomers of SAS-5: a trimeric coiled coil and a novel globular dimeric Implico domain. Disruption of either domain leads to centriole duplication failure in worm embryos, indicating that large SAS-5 assemblies are necessary for function in vivo.
中心粒是基于微管的细胞器,对细胞分裂、感知和运动至关重要。在秀丽隐杆线虫中,中心粒形成的起始特别需要蛋白质SAS-5和SAS-6,它们在真核生物进化过程中具有功能等同物。虽然SAS-6的分子结构及其在启动中心粒形成中的作用已得到充分了解,但SAS-5及其相关蛋白发挥功能的机制尚不清楚。在这里,我们结合生物物理和结构分析来揭示SAS-5的结构,并研究其在体内的功能意义。我们的工作表明,形成SAS-5的高阶寡聚体需要两个不同的自缔合结构域:一个三聚体卷曲螺旋和一个新的球状二聚体Implico结构域。破坏任一结构域都会导致线虫胚胎中的中心粒复制失败,这表明大型SAS-5组装体对于体内功能是必需的。