Vaquero-Garcia Jorge, Barrera Alejandro, Gazzara Matthew R, González-Vallinas Juan, Lahens Nicholas F, Hogenesch John B, Lynch Kristen W, Barash Yoseph
Department of Genetics, Perelman School of Medicine, University of Pennsylvania, Philadelphia, United States.
Department of Computer and Information Science, University of Pennsylvania, Philadelphia, United States.
Elife. 2016 Feb 1;5:e11752. doi: 10.7554/eLife.11752.
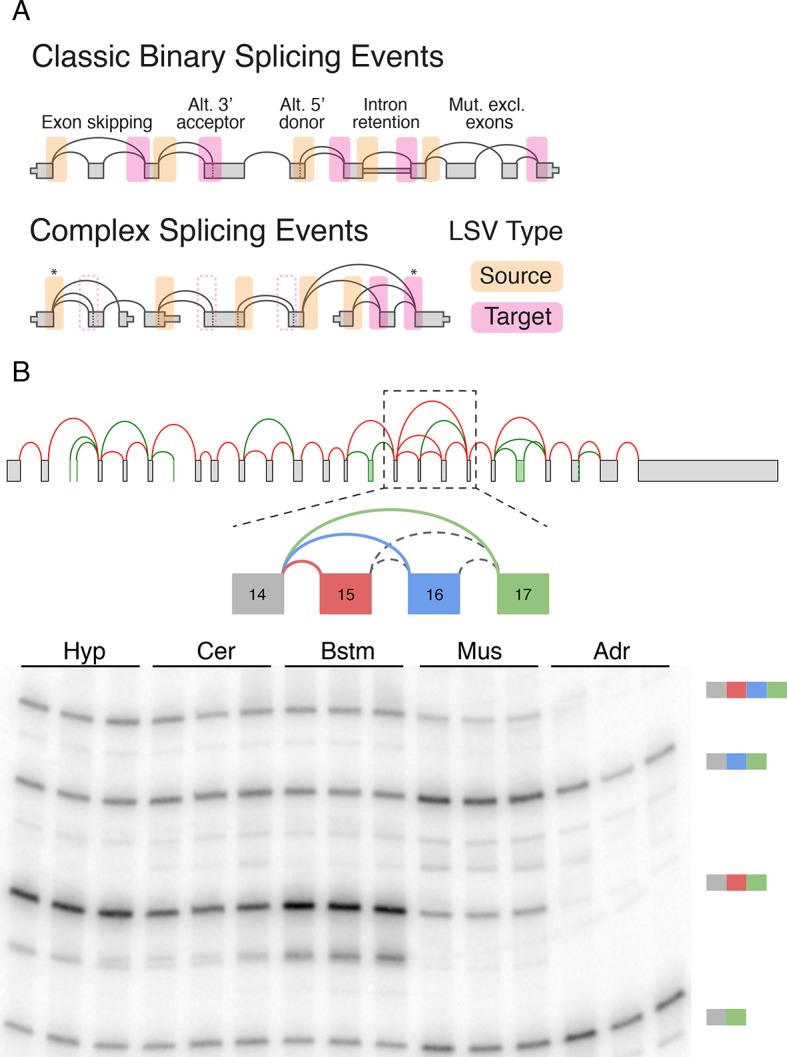
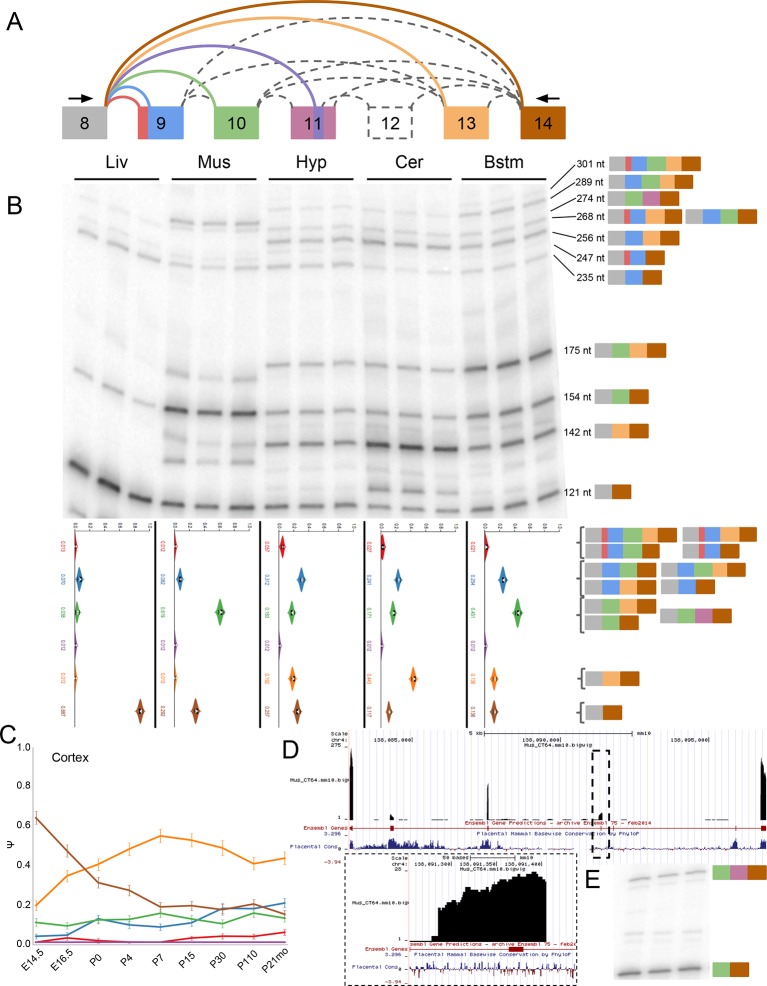
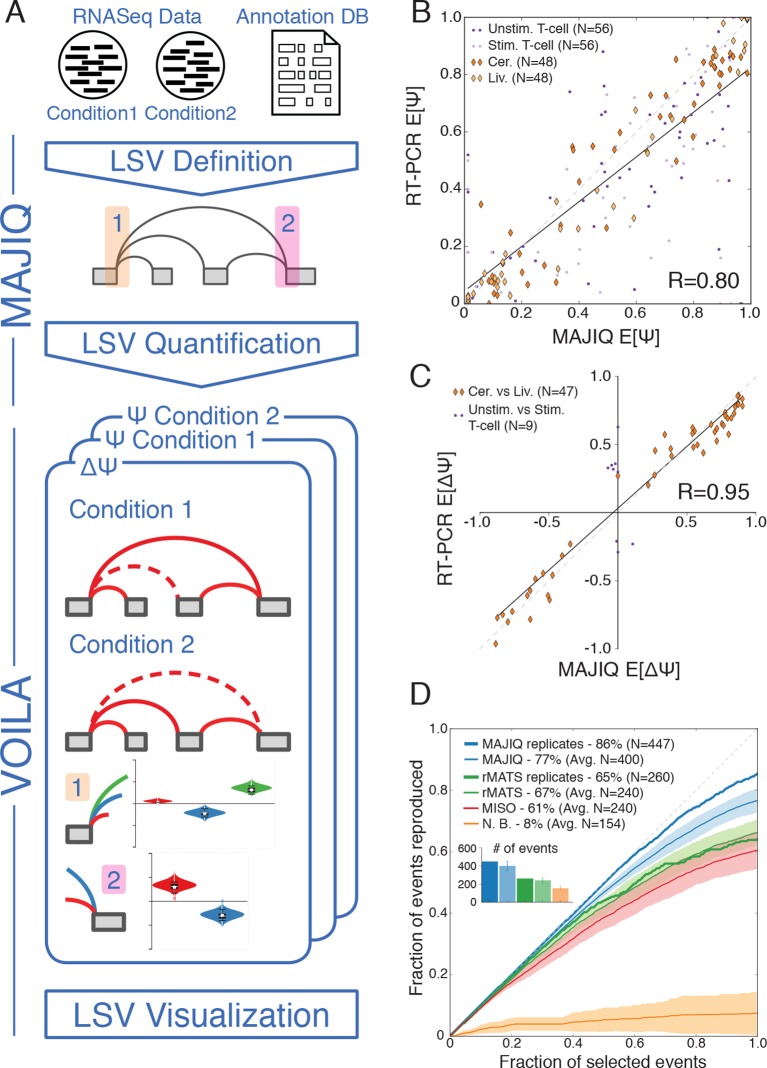
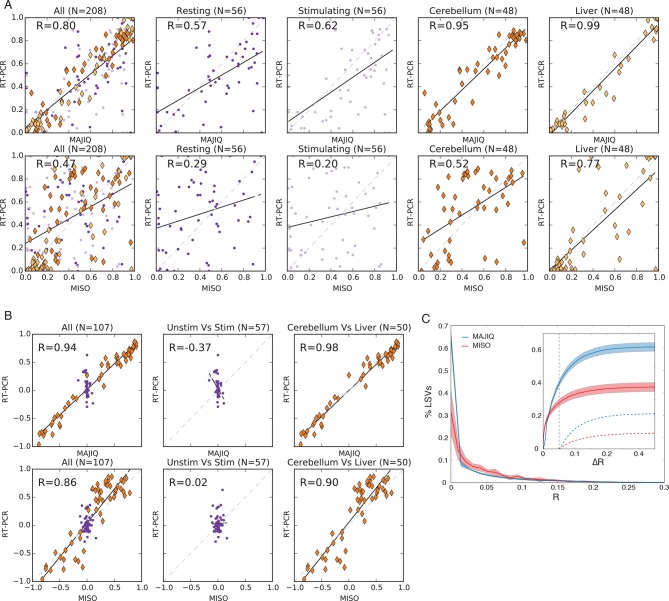
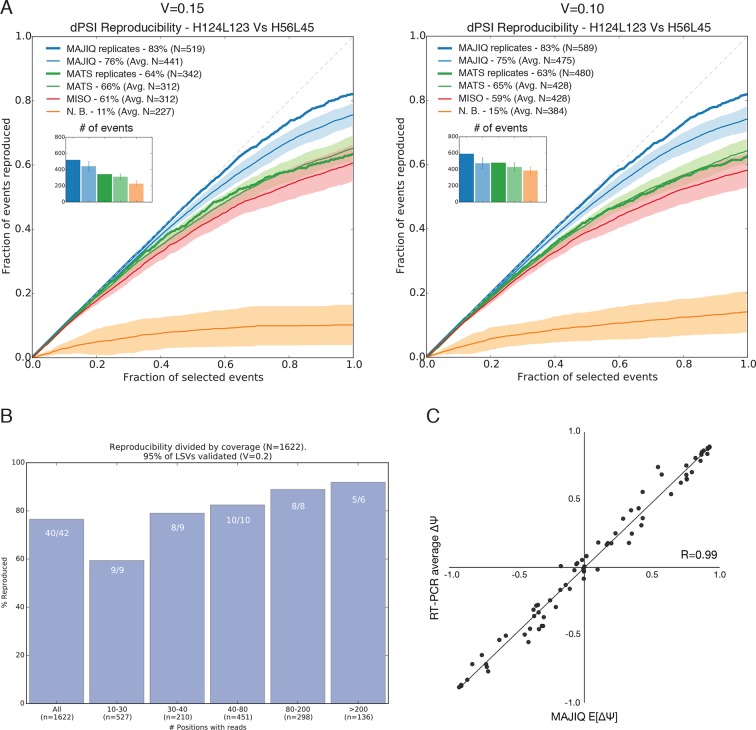
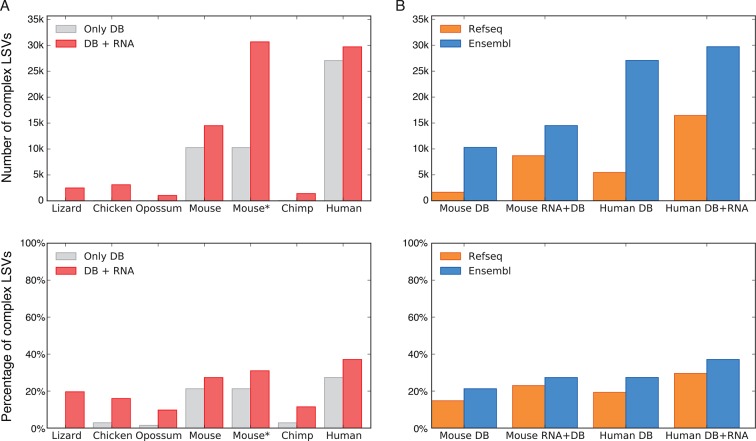
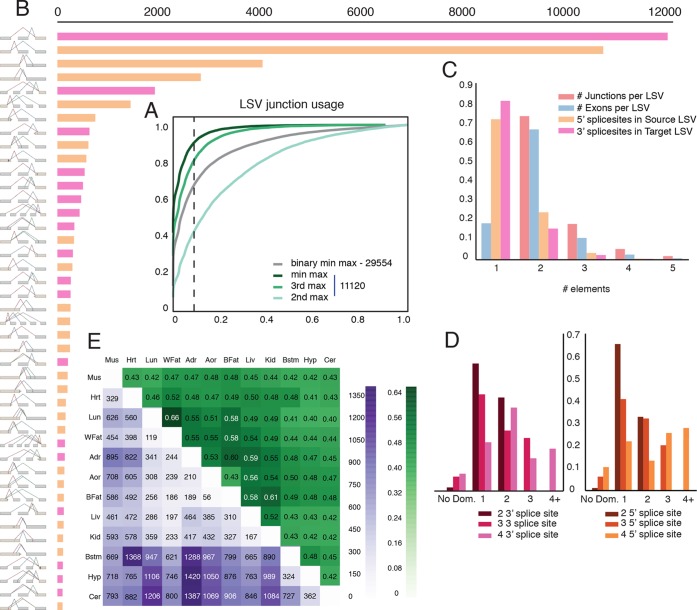
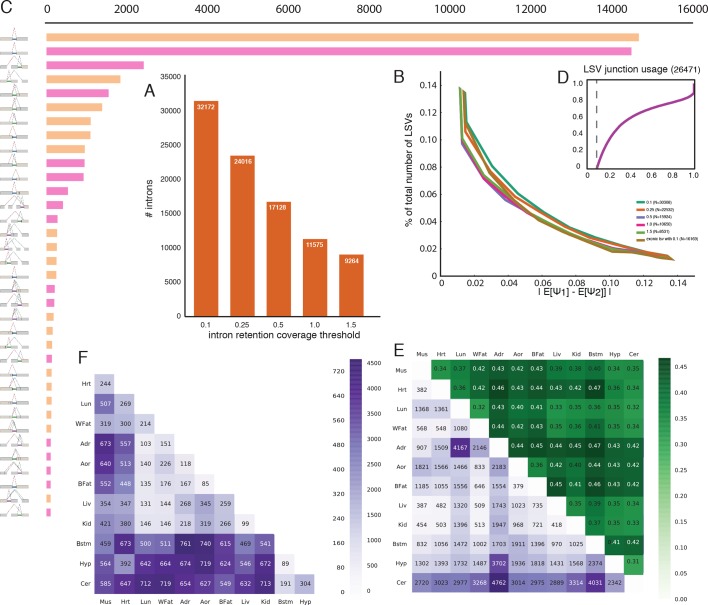
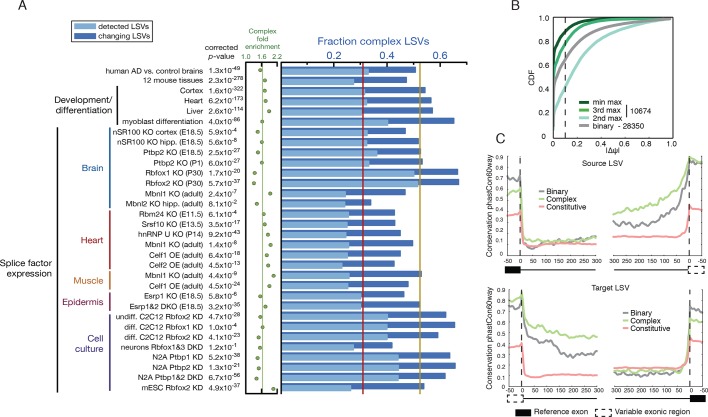
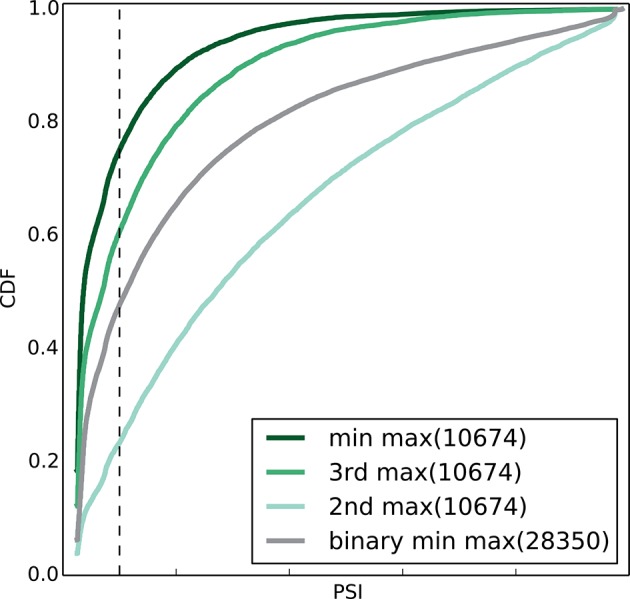
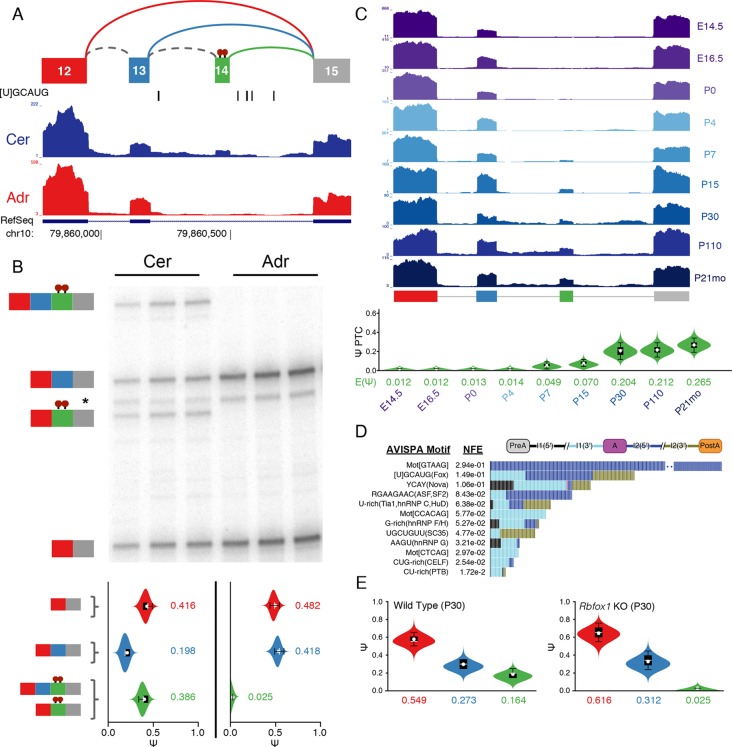
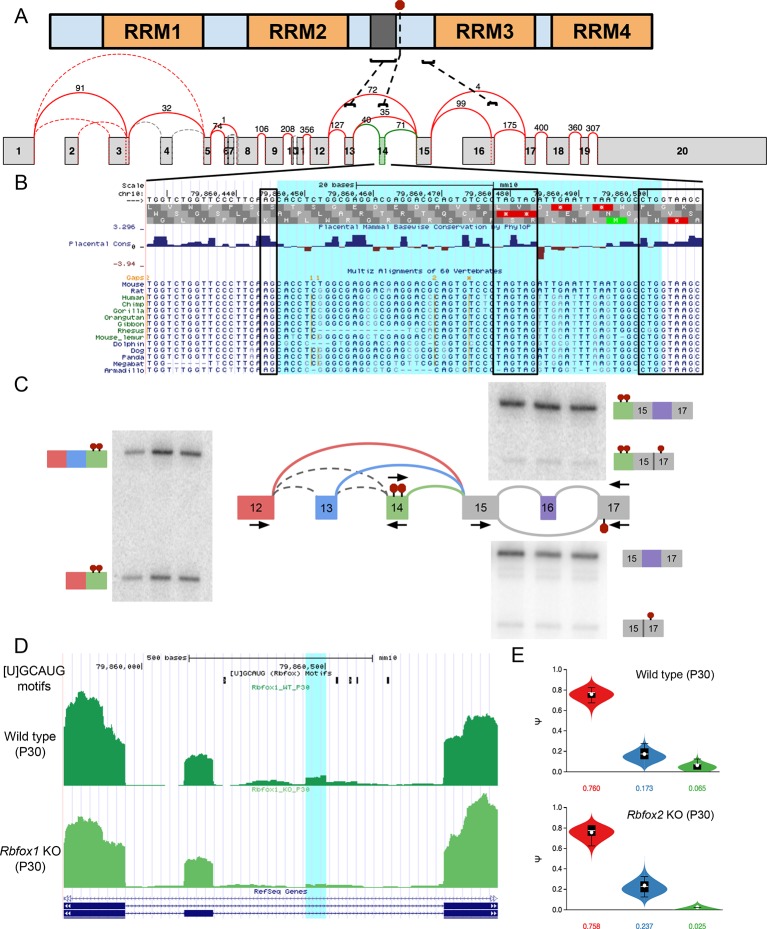
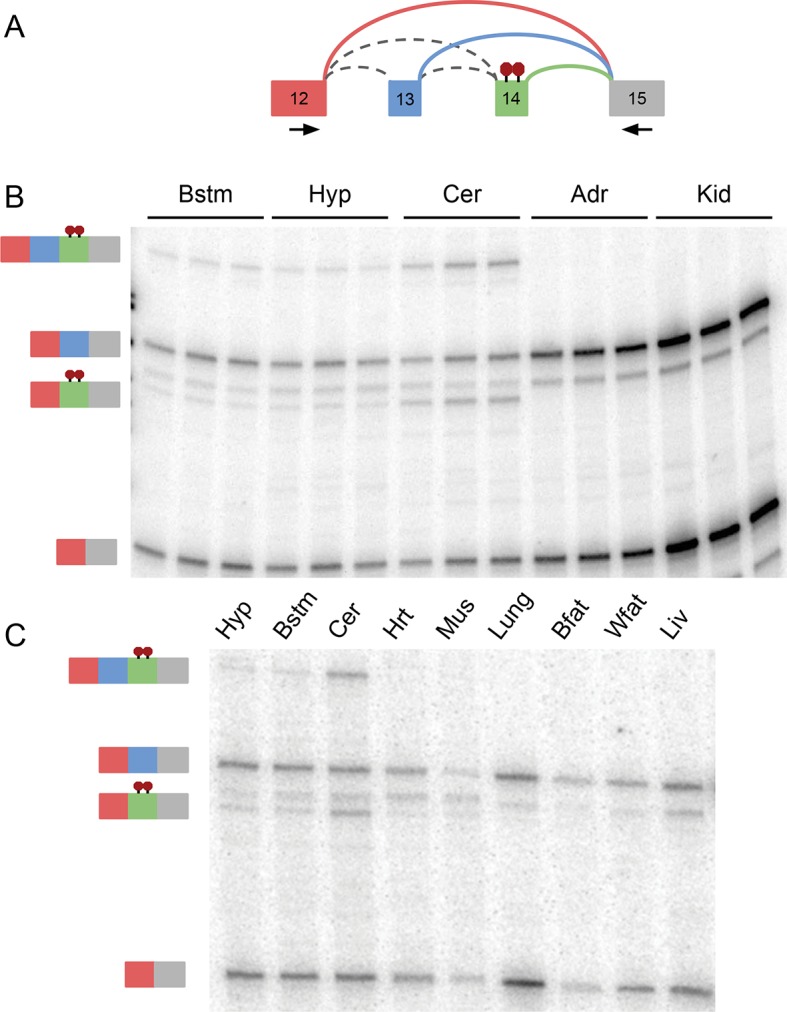
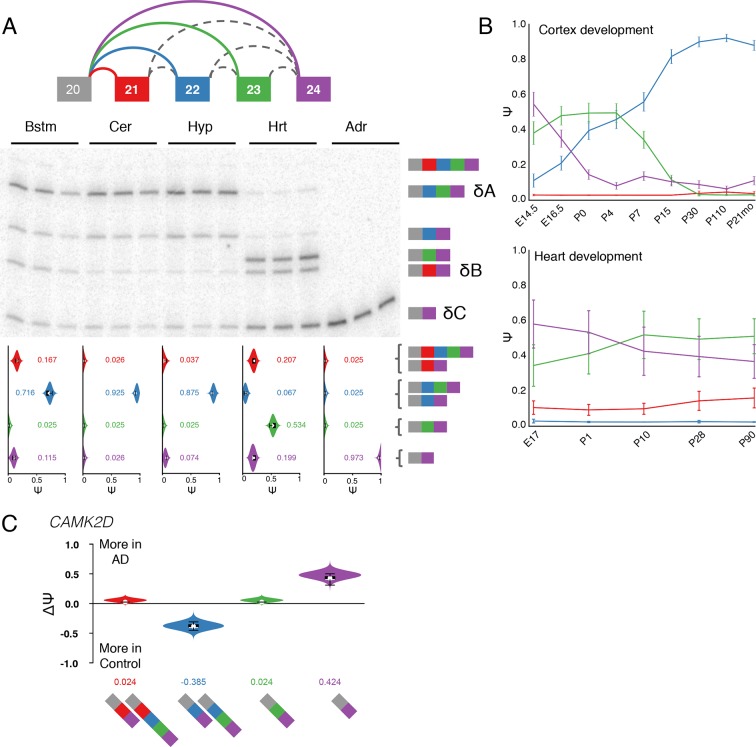
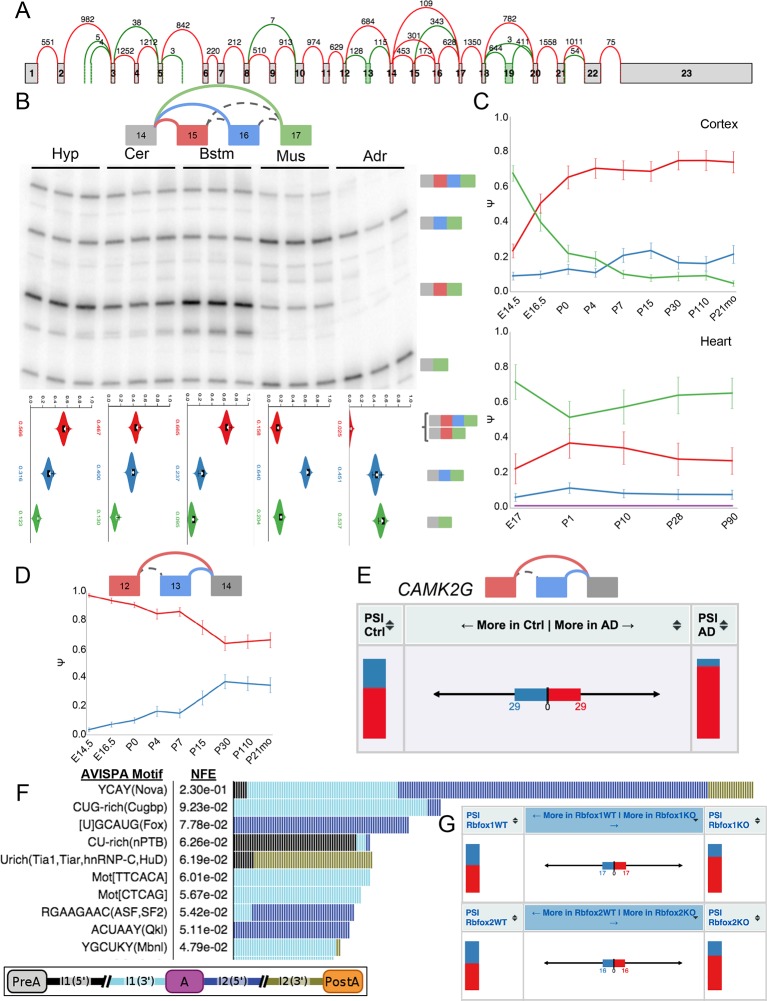
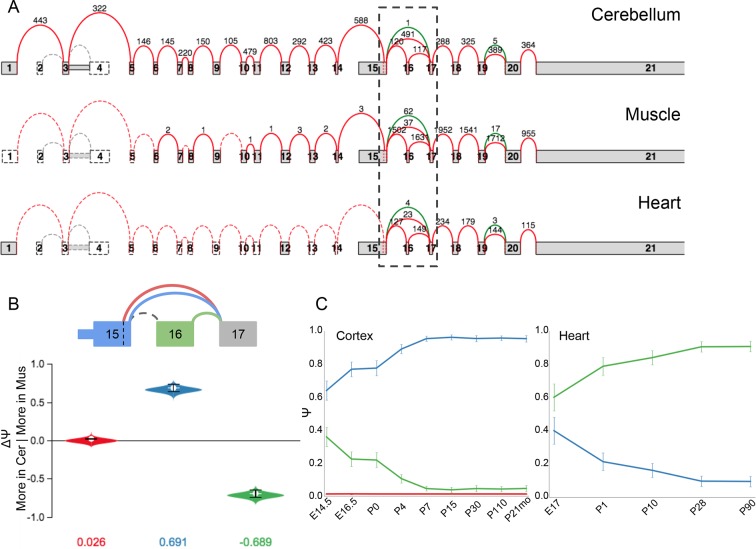
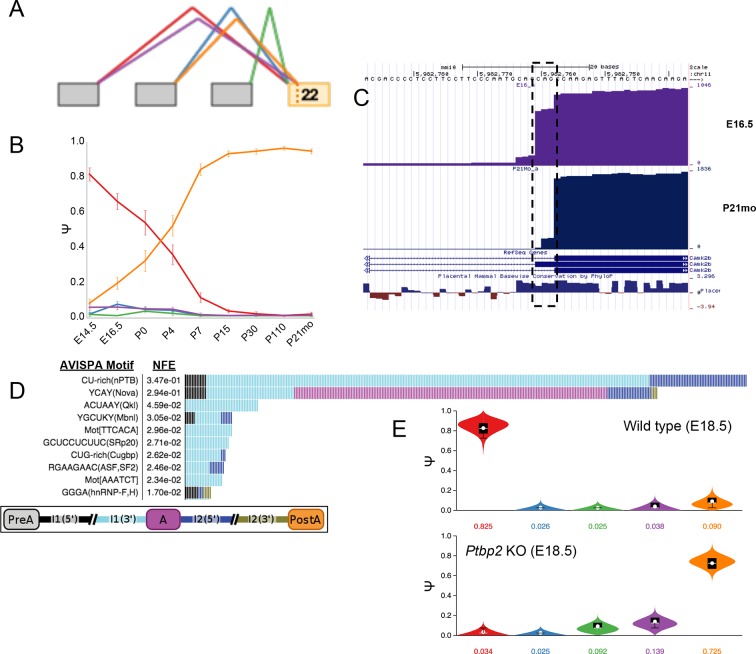
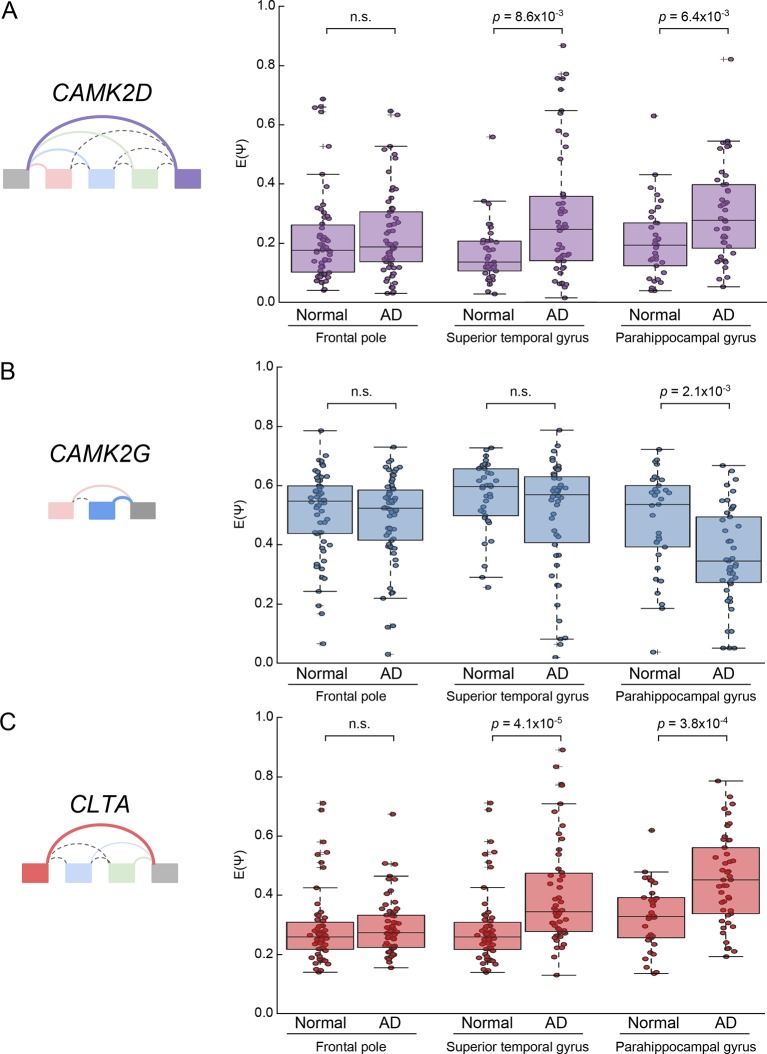
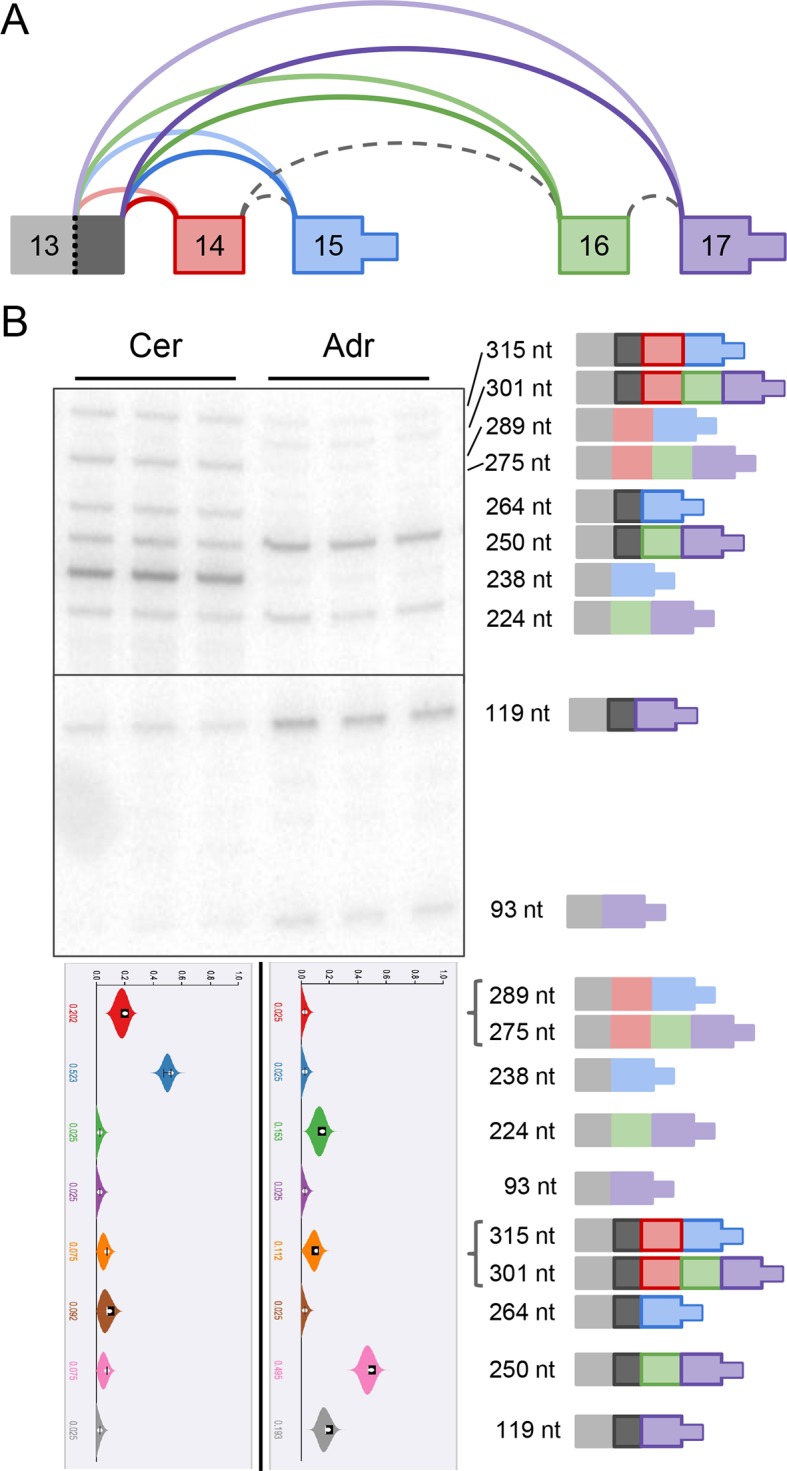
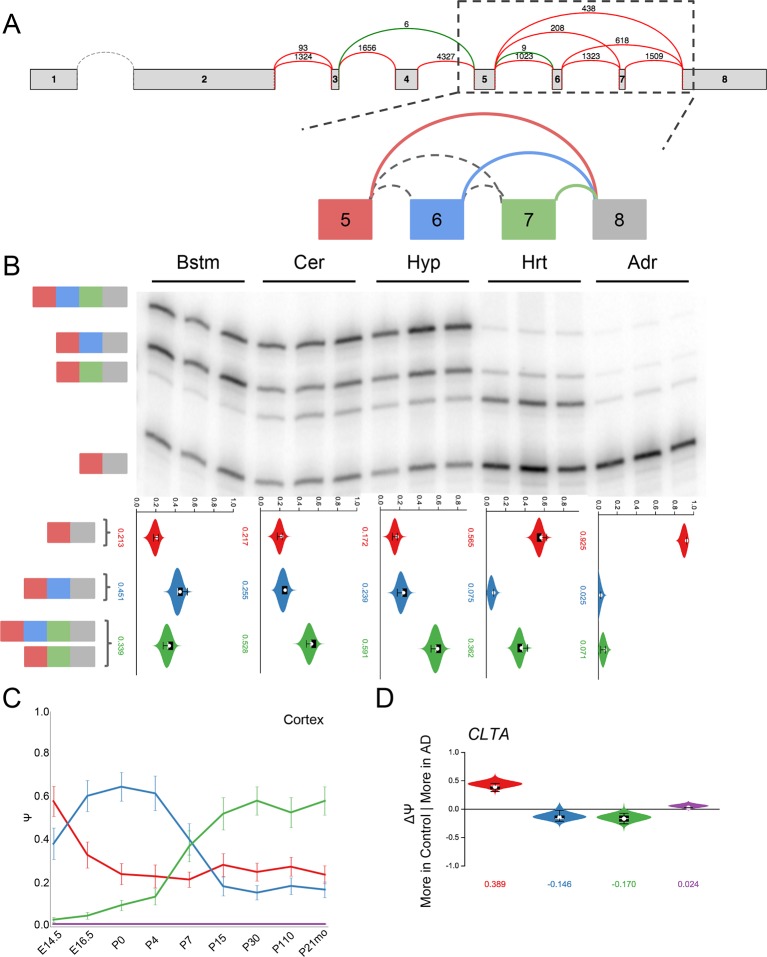
Alternative splicing (AS) can critically affect gene function and disease, yet mapping splicing variations remains a challenge. Here, we propose a new approach to define and quantify mRNA splicing in units of local splicing variations (LSVs). LSVs capture previously defined types of alternative splicing as well as more complex transcript variations. Building the first genome wide map of LSVs from twelve mouse tissues, we find complex LSVs constitute over 30% of tissue dependent transcript variations and affect specific protein families. We show the prevalence of complex LSVs is conserved in humans and identify hundreds of LSVs that are specific to brain subregions or altered in Alzheimer's patients. Amongst those are novel isoforms in the Camk2 family and a novel poison exon in Ptbp1, a key splice factor in neurogenesis. We anticipate the approach presented here will advance the ability to relate tissue-specific splice variation to genetic variation, phenotype, and disease.
可变剪接(Alternative splicing,AS)会对基因功能和疾病产生关键影响,但绘制剪接变异图谱仍然是一项挑战。在此,我们提出了一种新方法,以局部剪接变异(local splicing variations,LSVs)为单位来定义和量化mRNA剪接。LSVs涵盖了先前定义的可变剪接类型以及更复杂的转录本变异。通过构建来自12种小鼠组织的首张全基因组LSV图谱,我们发现复杂的LSVs占组织依赖性转录本变异的30%以上,并影响特定的蛋白质家族。我们表明复杂LSVs的普遍性在人类中是保守的,并鉴定出数百种特定于脑亚区域或在阿尔茨海默病患者中发生改变的LSVs。其中包括Camk2家族中的新型异构体以及Ptbp1(神经发生中的关键剪接因子)中的新型毒性外显子。我们预计本文提出的方法将提高将组织特异性剪接变异与遗传变异、表型和疾病联系起来的能力。