Jain Suvi, Sugawara Neal, Haber James E
Department of Biology and Rosenstiel Medical Center, Brandeis University, Waltham, Massachusetts, United States of America.
PLoS Genet. 2016 Apr 13;12(4):e1005976. doi: 10.1371/journal.pgen.1005976. eCollection 2016 Apr.
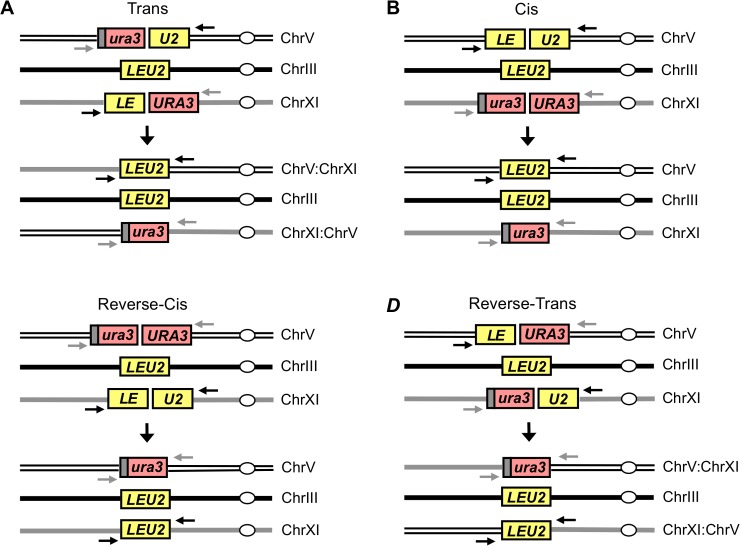
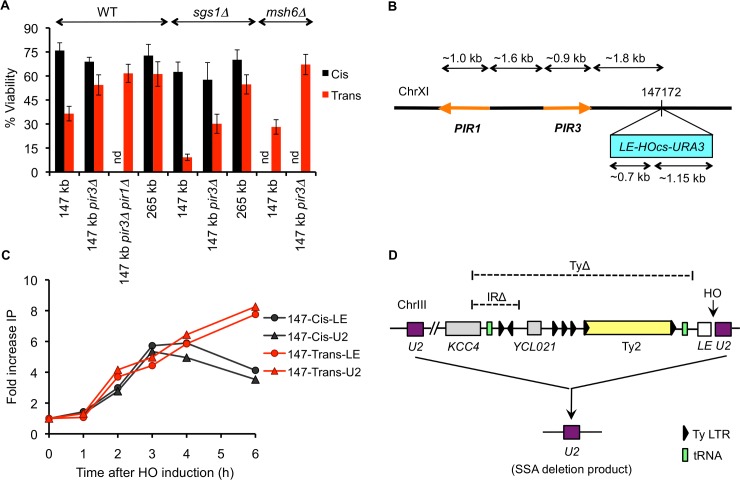
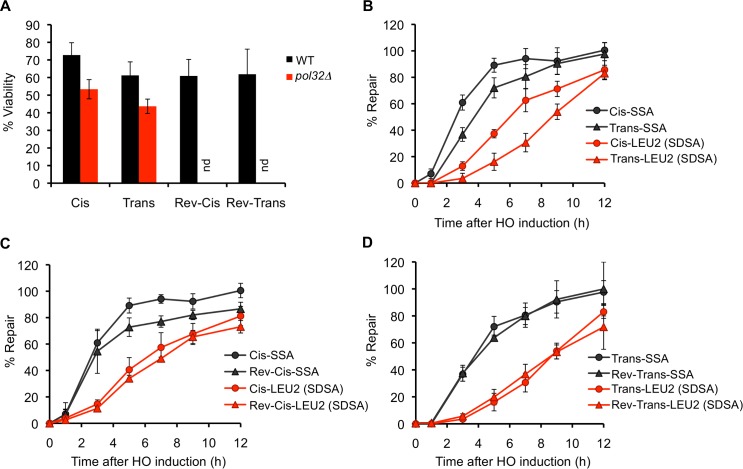
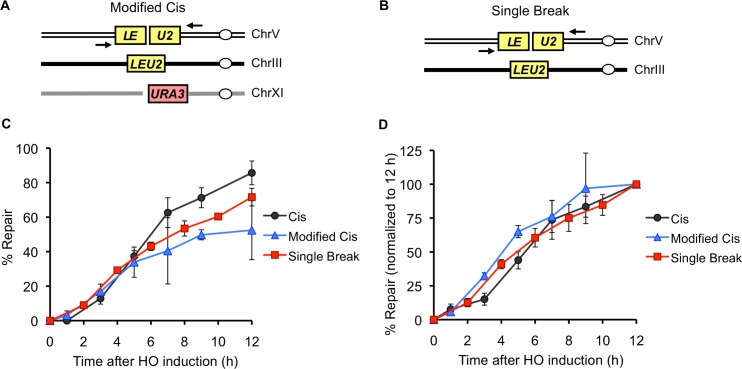
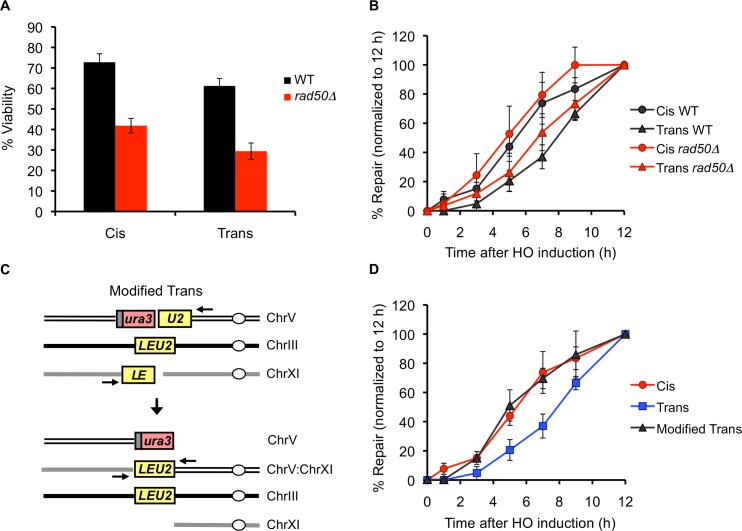
Correct repair of DNA double-strand breaks (DSBs) is critical for maintaining genome stability. Whereas gene conversion (GC)-mediated repair is mostly error-free, repair by break-induced replication (BIR) is associated with non-reciprocal translocations and loss of heterozygosity. We have previously shown that a Recombination Execution Checkpoint (REC) mediates this competition by preventing the BIR pathway from acting on DSBs that can be repaired by GC. Here, we asked if the REC can also determine whether the ends that are engaged in a GC-compatible configuration belong to the same break, since repair involving ends from different breaks will produce potentially deleterious translocations. We report that the kinetics of repair are markedly delayed when the two DSB ends that participate in GC belong to different DSBs (termed Trans) compared to the case when both DSB ends come from the same break (Cis). However, repair in Trans still occurs by GC rather than BIR, and the overall efficiency of repair is comparable. Hence, the REC is not sensitive to the "origin" of the DSB ends. When the homologous ends for GC are in Trans, the delay in repair appears to reflect their tethering to sequences on the other side of the DSB that themselves recombine with other genomic locations with which they share sequence homology. These data support previous observations that the two ends of a DSB are usually tethered to each other and that this tethering facilitates both ends encountering the same donor sequence. We also found that the presence of homeologous/repetitive sequences in the vicinity of a DSB can distract the DSB end from finding its bona fide homologous donor, and that inhibition of GC by such homeologous sequences is markedly increased upon deleting Sgs1 but not Msh6.
正确修复DNA双链断裂(DSB)对于维持基因组稳定性至关重要。基因转换(GC)介导的修复大多无差错,而断裂诱导复制(BIR)介导的修复则与非相互易位和杂合性丧失有关。我们之前已经表明,重组执行检查点(REC)通过阻止BIR途径作用于可由GC修复的DSB来介导这种竞争。在这里,我们询问REC是否还能确定处于GC兼容构型的末端是否属于同一断裂,因为涉及不同断裂末端的修复会产生潜在有害的易位。我们报告称,与两个DSB末端来自同一断裂(顺式)的情况相比,当参与GC的两个DSB末端属于不同DSB(称为反式)时,修复动力学明显延迟。然而,反式修复仍然通过GC而非BIR发生,并且总体修复效率相当。因此,REC对DSB末端的“起源”不敏感。当GC的同源末端处于反式时,修复延迟似乎反映了它们与DSB另一侧序列的连接,而这些序列本身会与它们共享序列同源性的其他基因组位置发生重组。这些数据支持了之前的观察结果,即DSB的两端通常相互连接,并且这种连接有助于两端遇到相同的供体序列。我们还发现,DSB附近存在同源/重复序列会使DSB末端难以找到其真正的同源供体,并且删除Sgs1而非Msh6会显著增加此类同源序列对GC的抑制作用。