Arenas-Salinas Mauricio, Vargas-Pérez Joaquín I, Morales Wladimir, Pinto Camilo, Muñoz-Díaz Pablo, Cornejo Fabián A, Pugin Benoit, Sandoval Juan M, Díaz-Vásquez Waldo A, Muñoz-Villagrán Claudia, Rodríguez-Rojas Fernanda, Morales Eduardo H, Vásquez Claudio C, Arenas Felipe A
Centro de Bioinformática y Simulación Molecular, Universidad de Talca Talca, Chile.
Departamento de Biología, Facultad de Química y Biología, Universidad de Santiago de Chile Santiago, Chile.
Front Microbiol. 2016 Jul 26;7:1160. doi: 10.3389/fmicb.2016.01160. eCollection 2016.
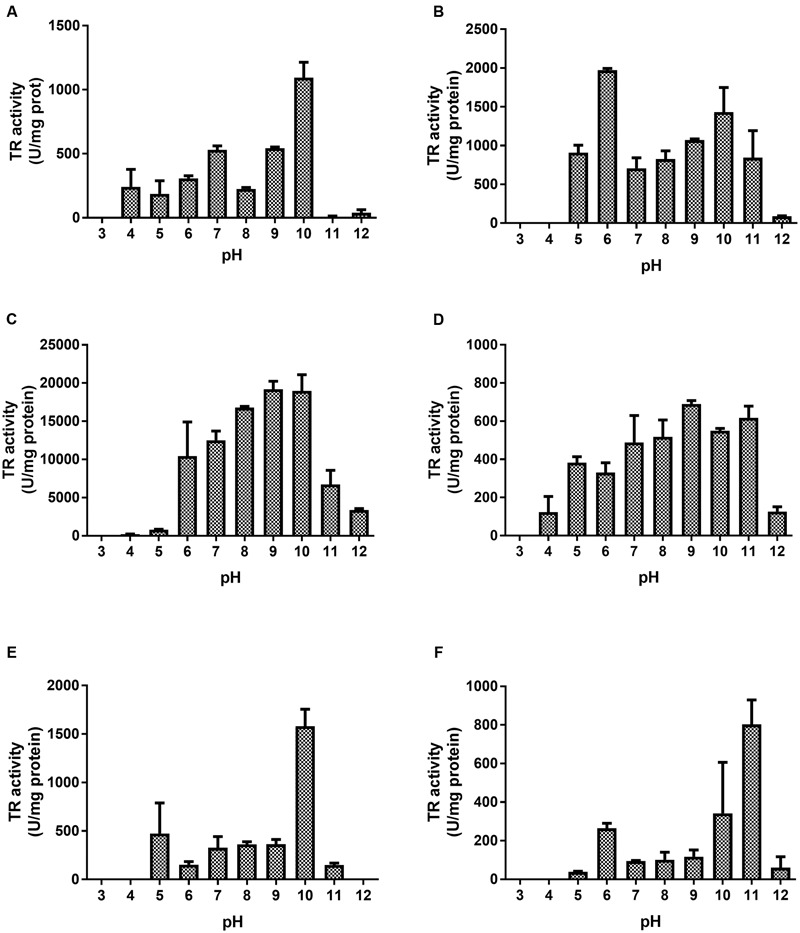
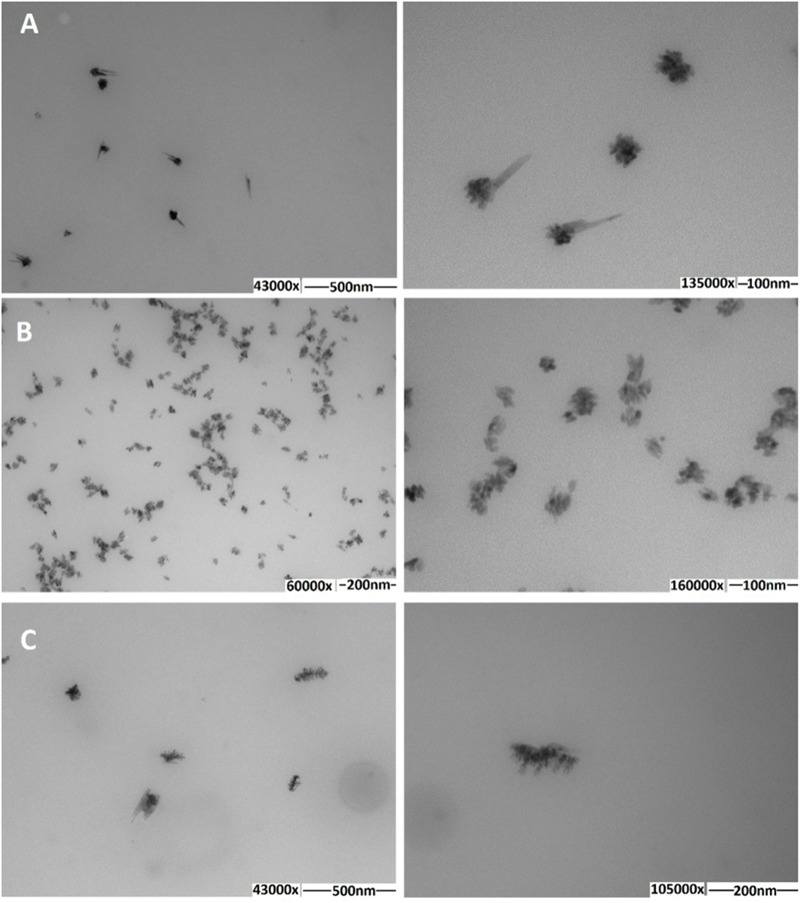
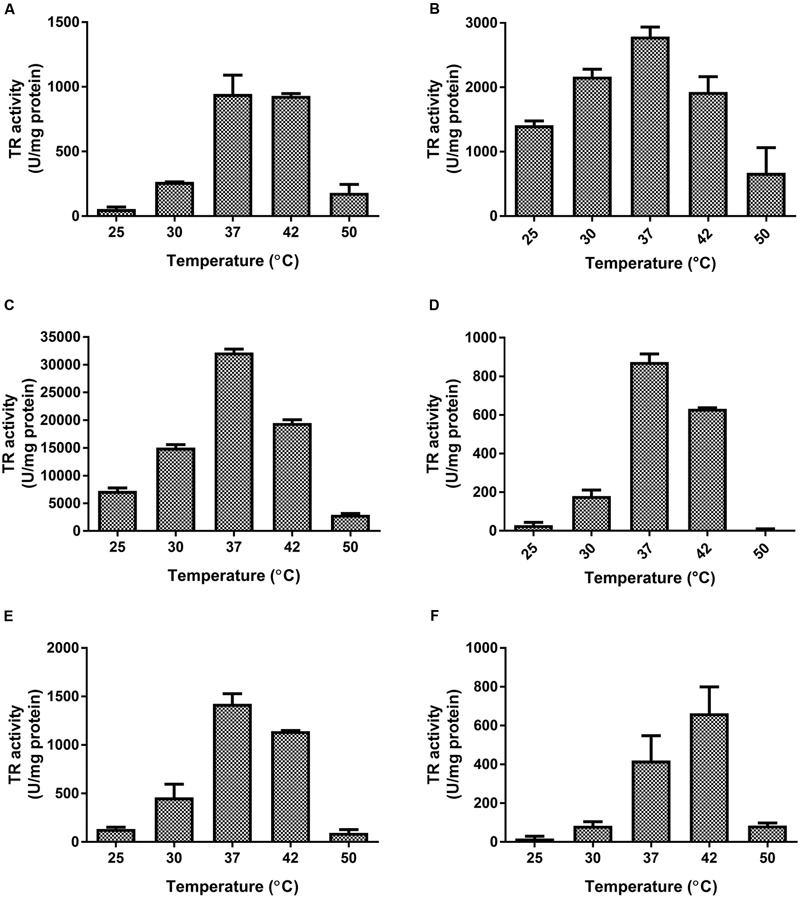
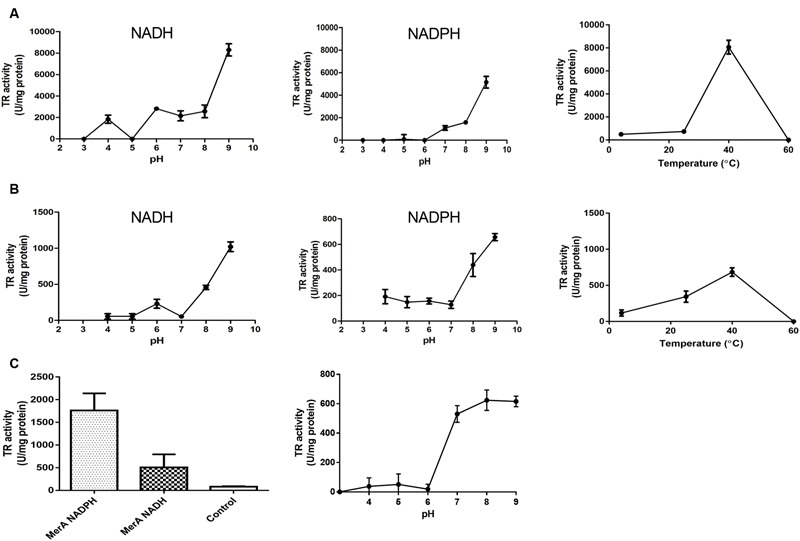
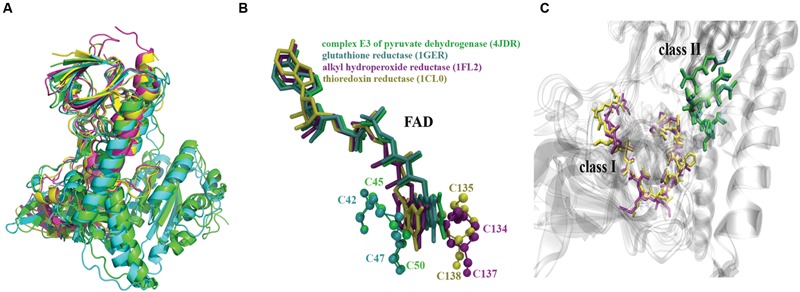
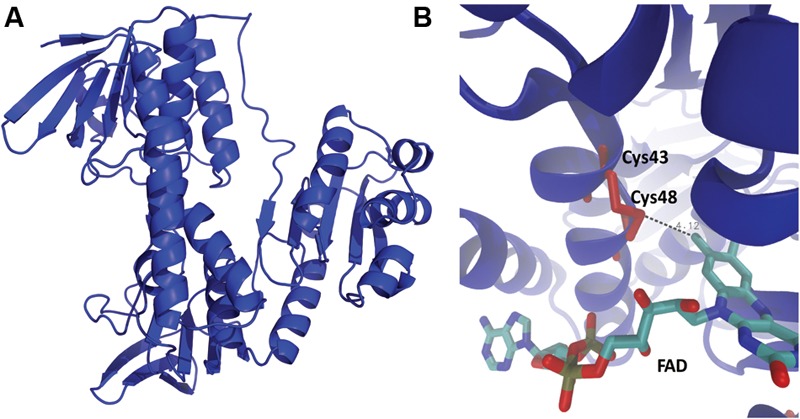
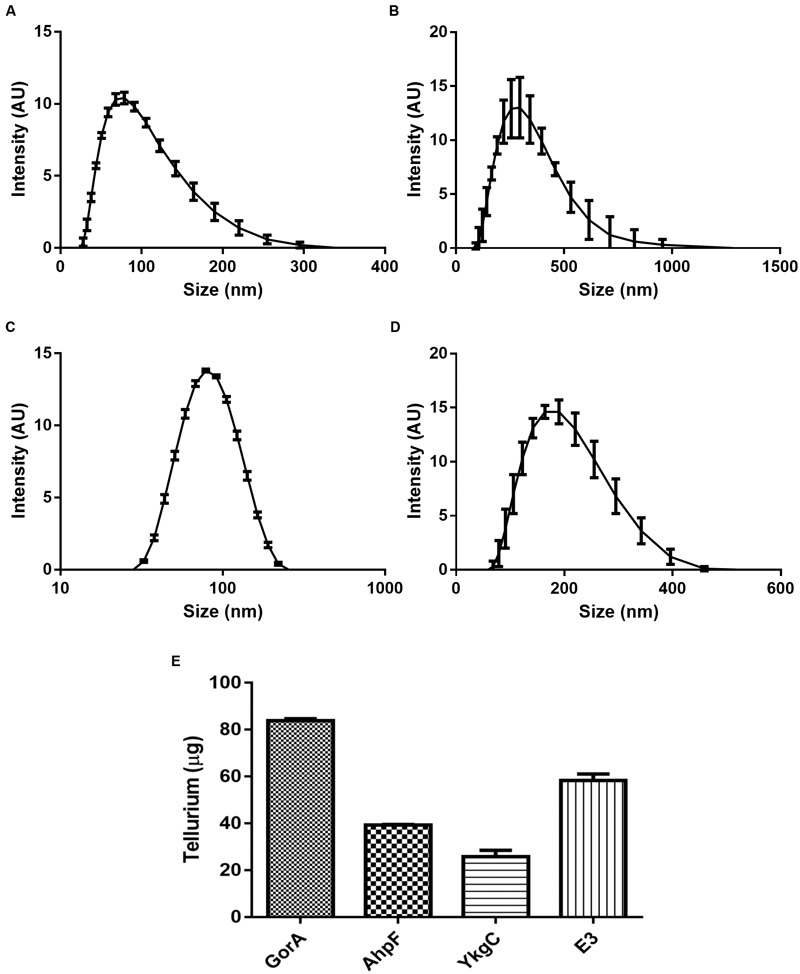
The tellurium oxyanion tellurite (TeO3 (2-)) is extremely harmful for most organisms. It has been suggested that a potential bacterial tellurite resistance mechanism would consist of an enzymatic, NAD(P)H-dependent, reduction to the less toxic form elemental tellurium (Te(0)). To date, a number of enzymes such as catalase, type II NADH dehydrogenase and terminal oxidases from the electron transport chain, nitrate reductases, and dihydrolipoamide dehydrogenase (E3), among others, have been shown to display tellurite-reducing activity. This activity is generically referred to as tellurite reductase (TR). Bioinformatic data resting on some of the abovementioned enzymes enabled the identification of common structures involved in tellurite reduction including vicinal catalytic cysteine residues and the FAD/NAD(P)(+)-binding domain, which is characteristic of some flavoproteins. Along this line, thioredoxin reductase (TrxB), alkyl hydroperoxide reductase (AhpF), glutathione reductase (GorA), mercuric reductase (MerA), NADH: flavorubredoxin reductase (NorW), dihydrolipoamide dehydrogenase, and the putative oxidoreductase YkgC from Escherichia coli or environmental bacteria were purified and assessed for TR activity. All of them displayed in vitro TR activity at the expense of NADH or NADPH oxidation. In general, optimal reducing conditions occurred around pH 9-10 and 37°C. Enzymes exhibiting strong TR activity produced Te-containing nanostructures (TeNS). While GorA and AhpF generated TeNS of 75 nm average diameter, E3 and YkgC produced larger structures (>100 nm). Electron-dense structures were observed in cells over-expressing genes encoding TrxB, GorA, and YkgC.
氧碲酸根离子亚碲酸盐(TeO3 (2-))对大多数生物体极具危害。有人提出,一种潜在的细菌抗亚碲酸盐机制可能包括一种酶促反应,即依赖烟酰胺腺嘌呤二核苷酸(磷酸)(NAD(P)H)将其还原为毒性较小的元素碲(Te(0))。迄今为止,已证明多种酶,如过氧化氢酶、电子传递链中的II型NADH脱氢酶和末端氧化酶、硝酸还原酶以及二氢硫辛酰胺脱氢酶(E3)等,均具有亚碲酸盐还原活性。这种活性通常被称为亚碲酸盐还原酶(TR)。基于上述部分酶的生物信息学数据,得以鉴定出参与亚碲酸盐还原的常见结构,包括相邻的催化半胱氨酸残基以及FAD/NAD(P)(+)结合结构域,这是一些黄素蛋白的特征。据此,对来自大肠杆菌或环境细菌的硫氧还蛋白还原酶(TrxB)、烷基过氧化氢还原酶(AhpF)、谷胱甘肽还原酶(GorA)、汞还原酶(MerA)、NADH:黄素铁氧化还原蛋白还原酶(NorW)、二氢硫辛酰胺脱氢酶以及推定的氧化还原酶YkgC进行了纯化,并评估其TR活性。它们均以NADH或NADPH氧化为代价,在体外表现出TR活性。一般而言,最佳还原条件出现在pH 9 - 10和37°C左右。表现出强TR活性的酶产生了含碲纳米结构(TeNS)。虽然GorA和AhpF产生的TeNS平均直径为75 nm,但E3和YkgC产生的结构更大(>100 nm)。在过表达编码TrxB、GorA和YkgC基因的细胞中观察到了电子致密结构。