Keenan William Thomas, Rupp Alan C, Ross Rachel A, Somasundaram Preethi, Hiriyanna Suja, Wu Zhijian, Badea Tudor C, Robinson Phyllis R, Lowell Bradford B, Hattar Samer S
Department of Biology, Johns Hopkins University, Baltimore, United States.
Department of Psychiatry, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, United States.
Elife. 2016 Sep 26;5:e15392. doi: 10.7554/eLife.15392.
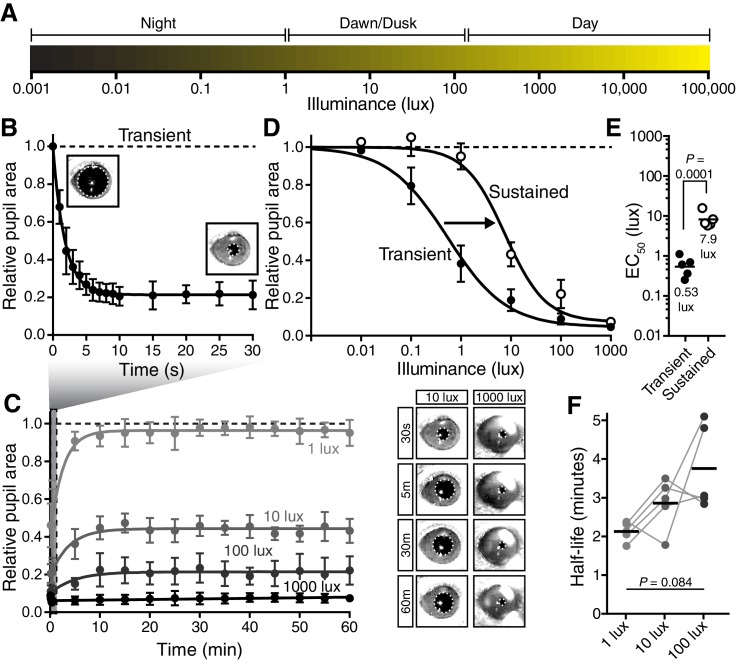
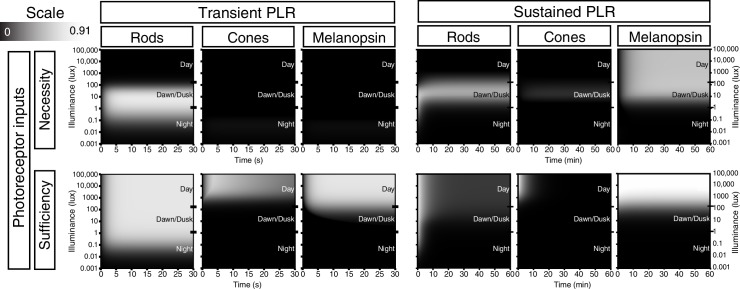
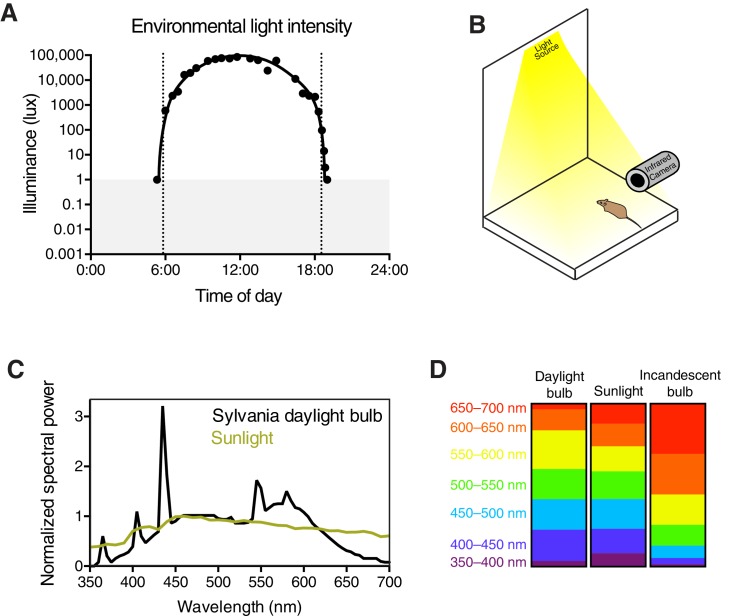
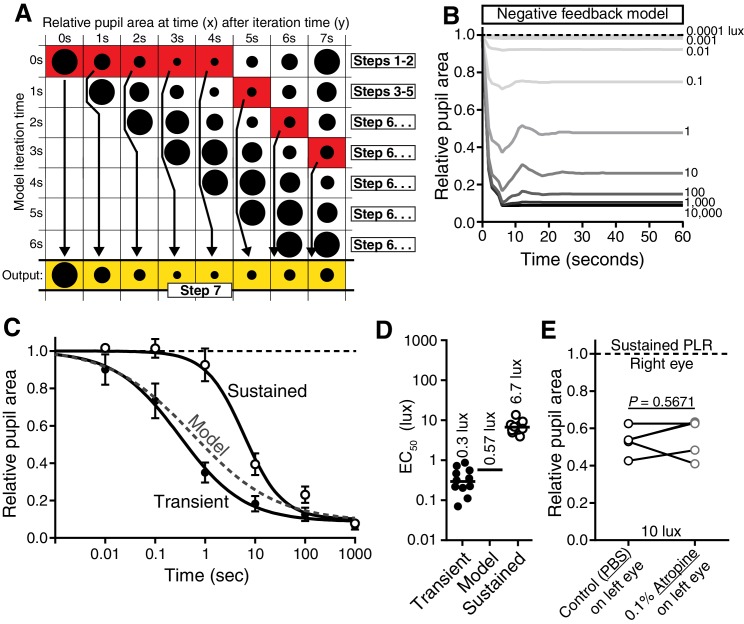
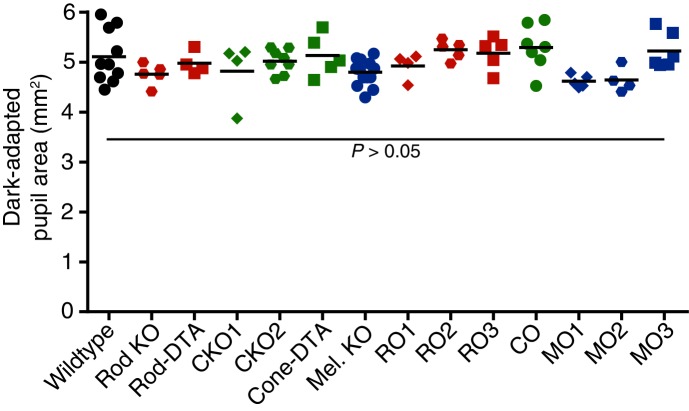
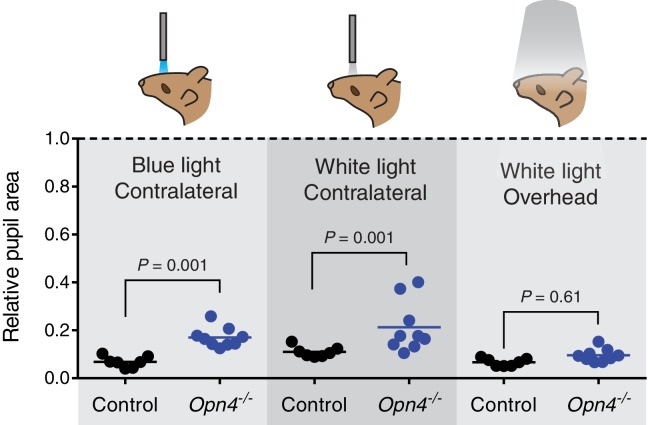
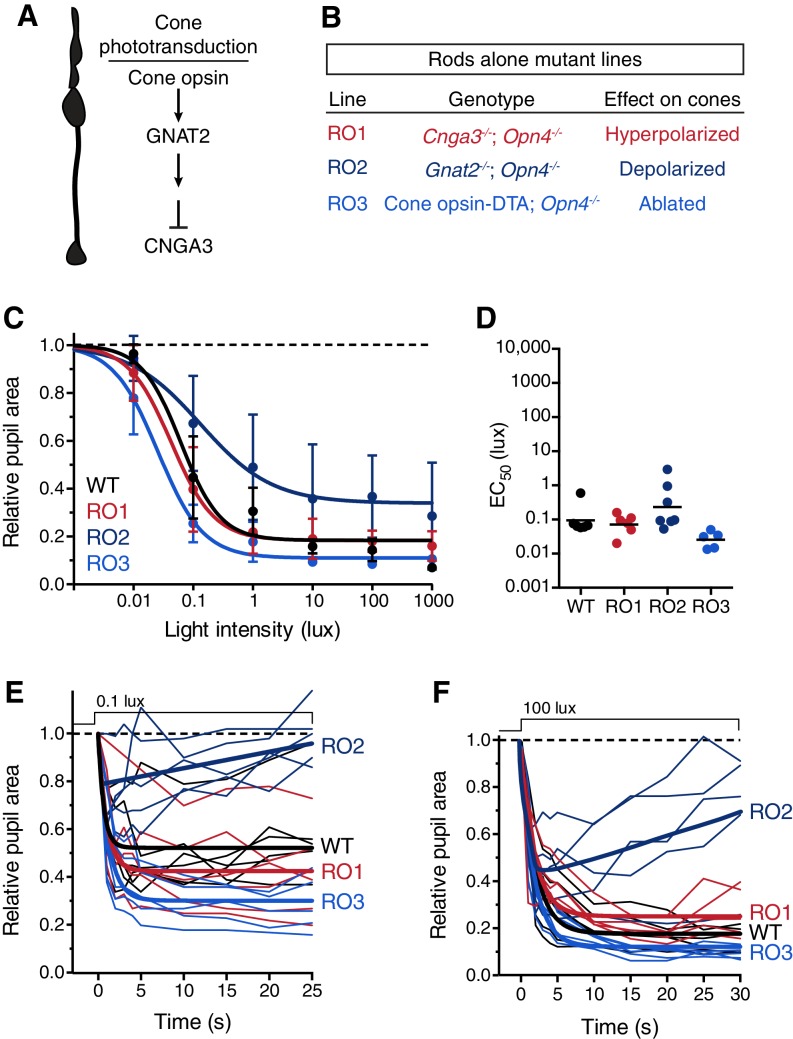
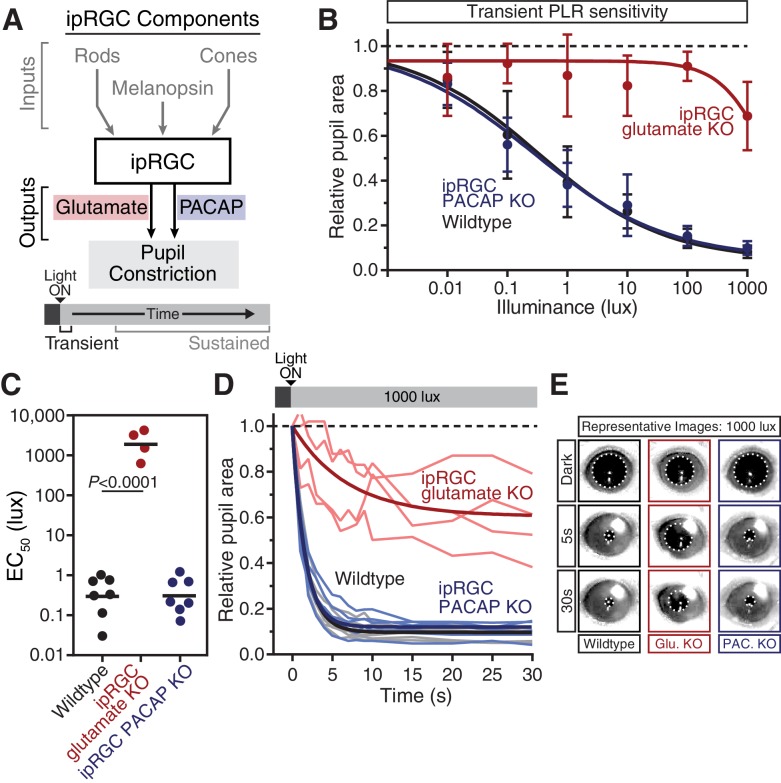
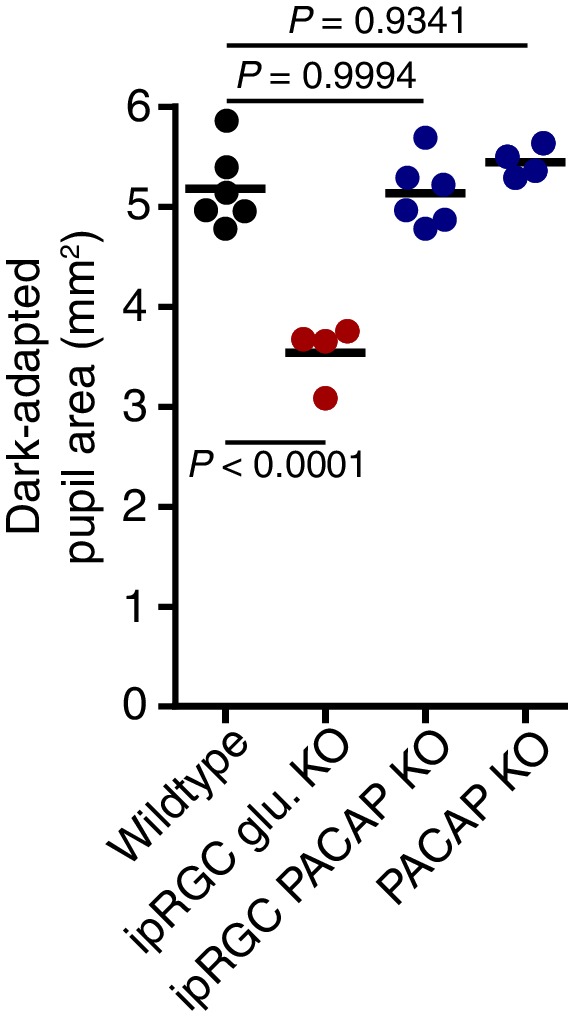
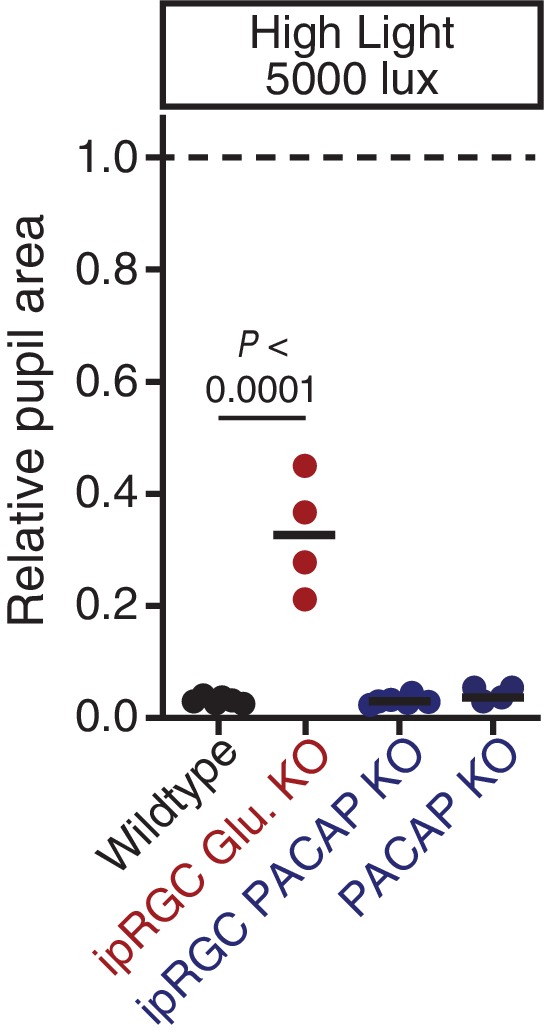
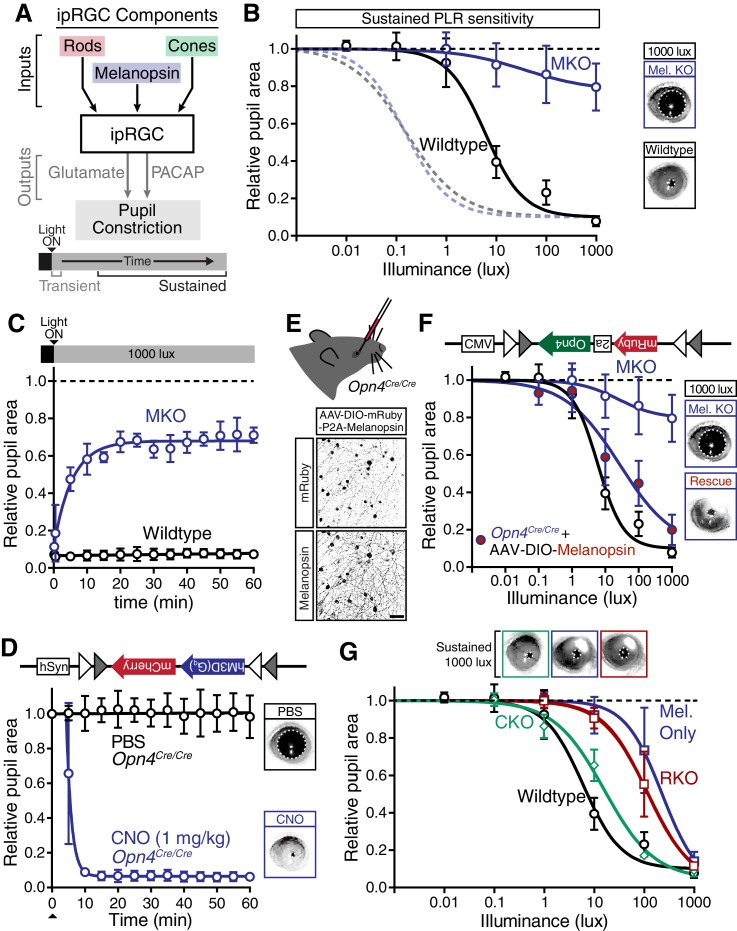
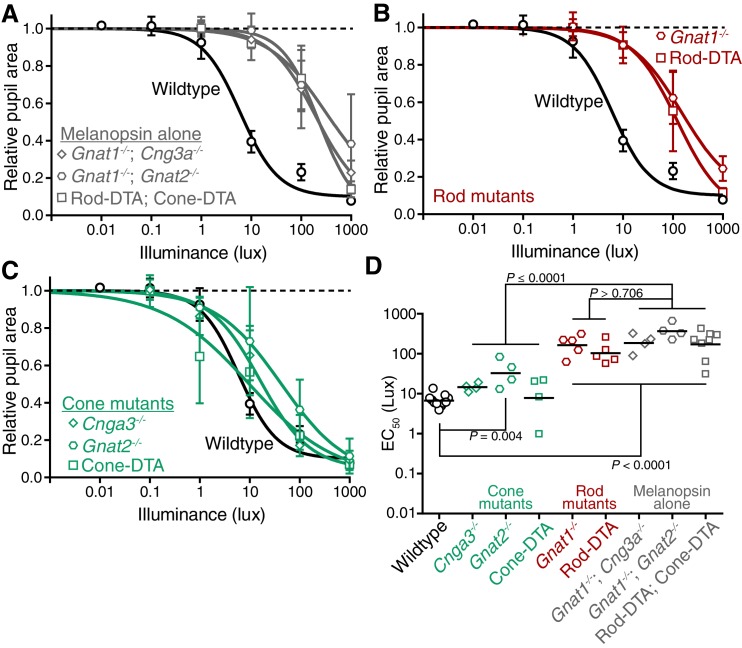
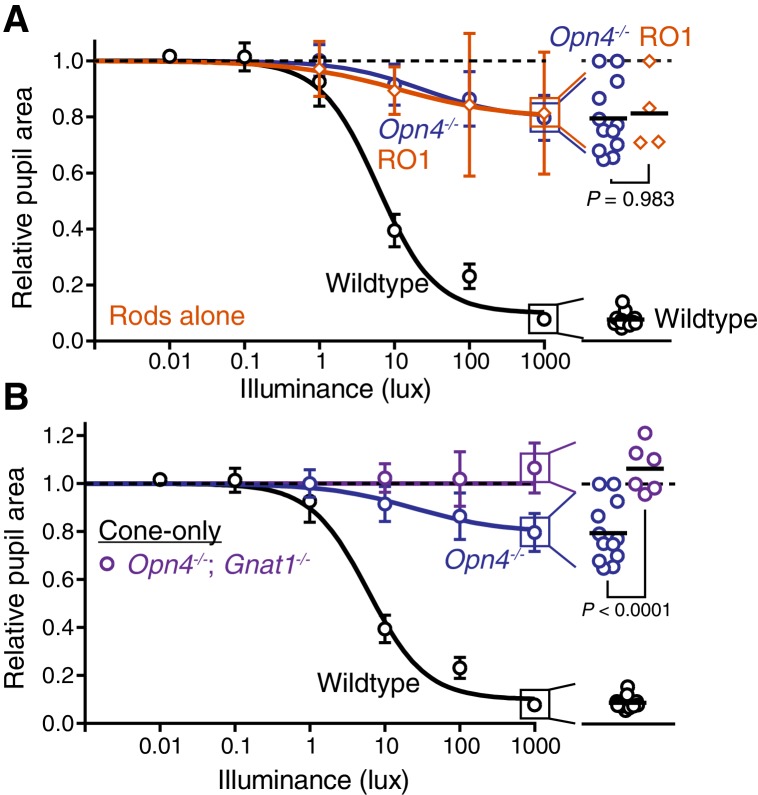
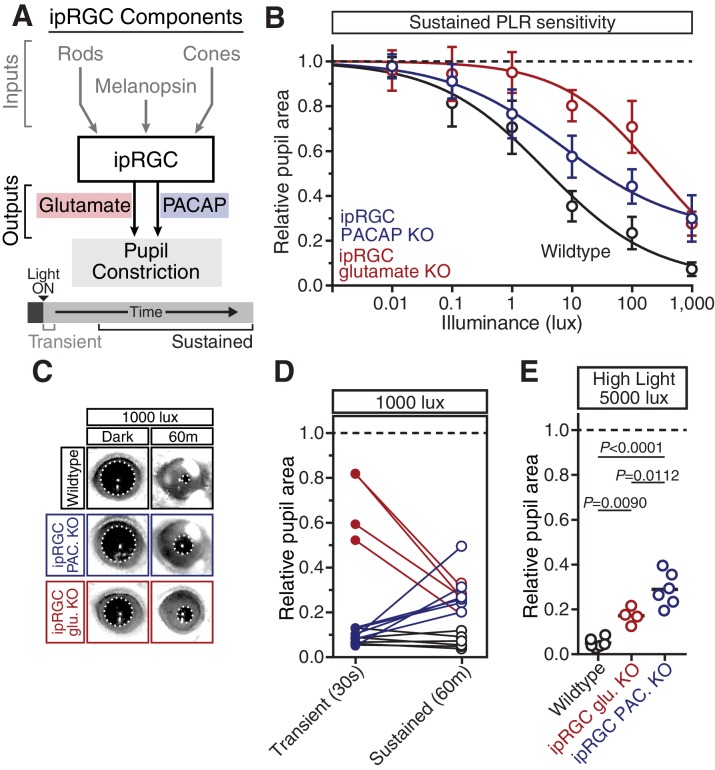
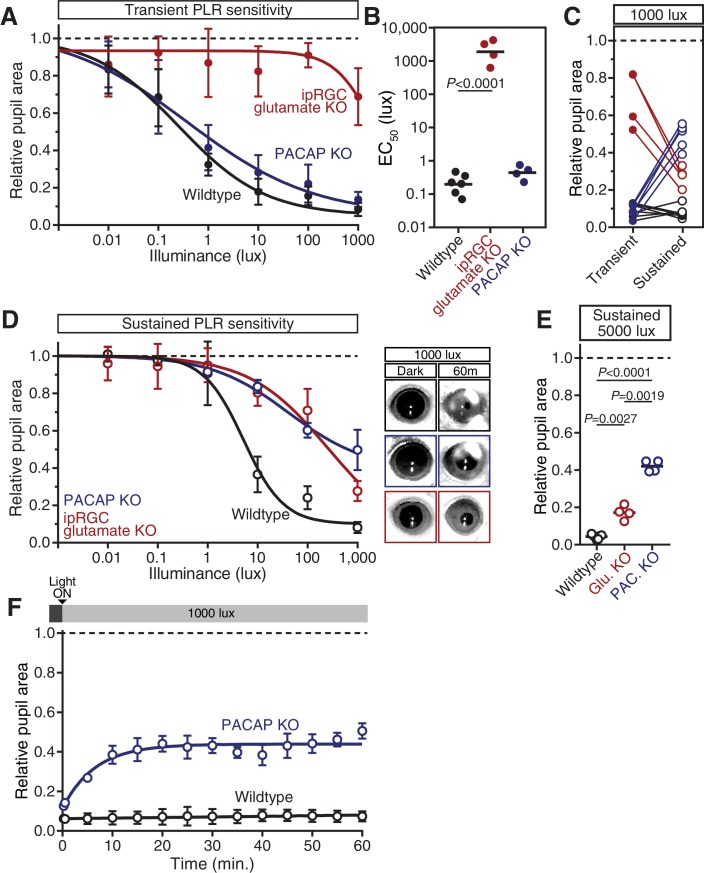
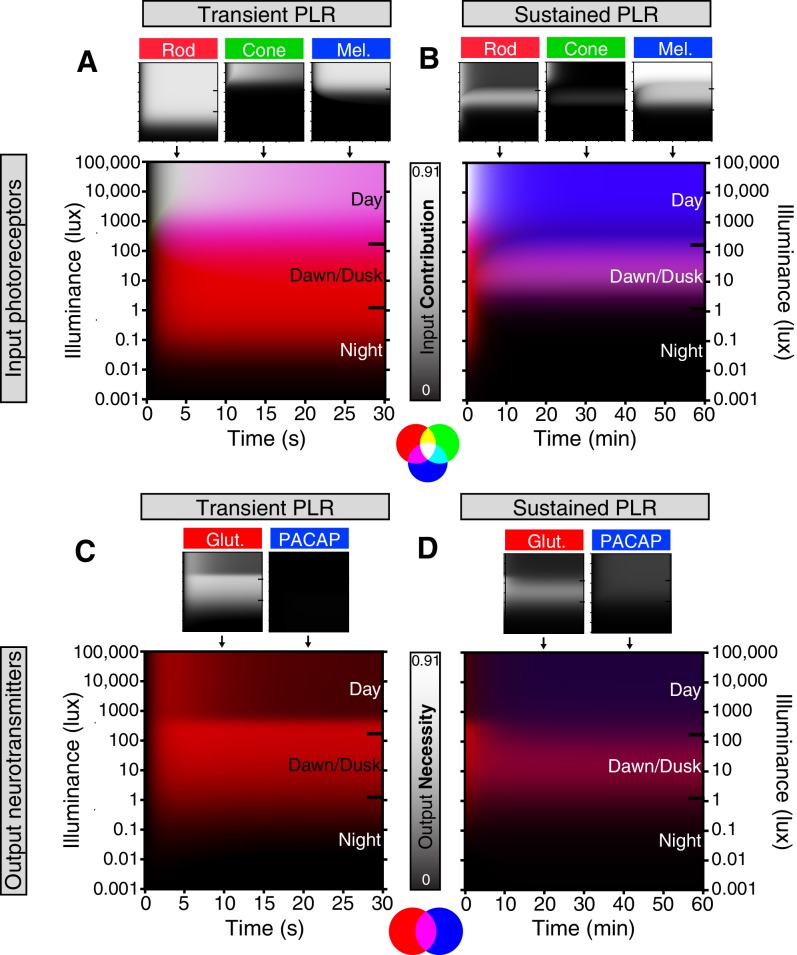
Rapid and stable control of pupil size in response to light is critical for vision, but the neural coding mechanisms remain unclear. Here, we investigated the neural basis of pupil control by monitoring pupil size across time while manipulating each photoreceptor input or neurotransmitter output of intrinsically photosensitive retinal ganglion cells (ipRGCs), a critical relay in the control of pupil size. We show that transient and sustained pupil responses are mediated by distinct photoreceptors and neurotransmitters. Transient responses utilize input from rod photoreceptors and output by the classical neurotransmitter glutamate, but adapt within minutes. In contrast, sustained responses are dominated by non-conventional signaling mechanisms: melanopsin phototransduction in ipRGCs and output by the neuropeptide PACAP, which provide stable pupil maintenance across the day. These results highlight a temporal switch in the coding mechanisms of a neural circuit to support proper behavioral dynamics.
对光作出快速且稳定的瞳孔大小控制对视觉至关重要,但神经编码机制仍不清楚。在此,我们通过在操纵内在光敏性视网膜神经节细胞(ipRGCs)的每个光感受器输入或神经递质输出时跨时间监测瞳孔大小,来研究瞳孔控制的神经基础,ipRGCs是瞳孔大小控制中的关键中继。我们表明,瞬态和持续的瞳孔反应由不同的光感受器和神经递质介导。瞬态反应利用视杆光感受器的输入,并通过经典神经递质谷氨酸输出,但在数分钟内会适应。相比之下,持续反应由非常规信号机制主导:ipRGCs中的黑视蛋白光转导以及神经肽垂体腺苷酸环化酶激活肽(PACAP)的输出,这在一整天内提供稳定的瞳孔维持。这些结果突出了神经回路编码机制中的一种时间转换,以支持适当的行为动态。