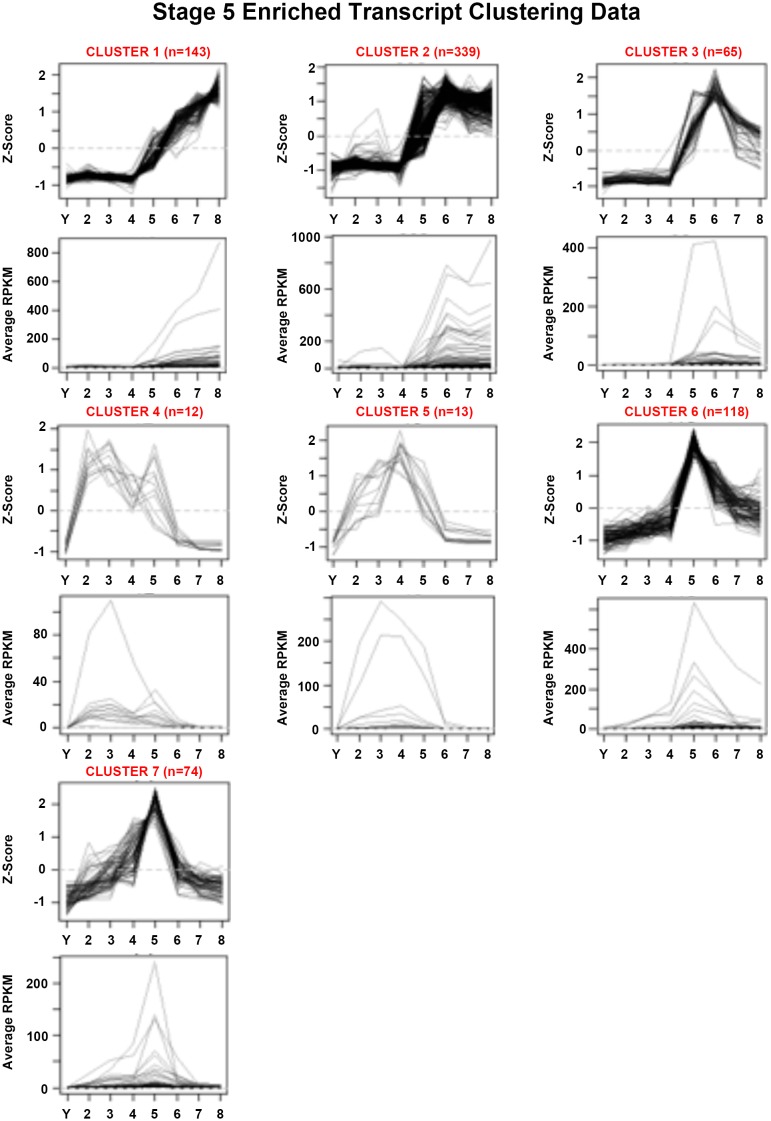
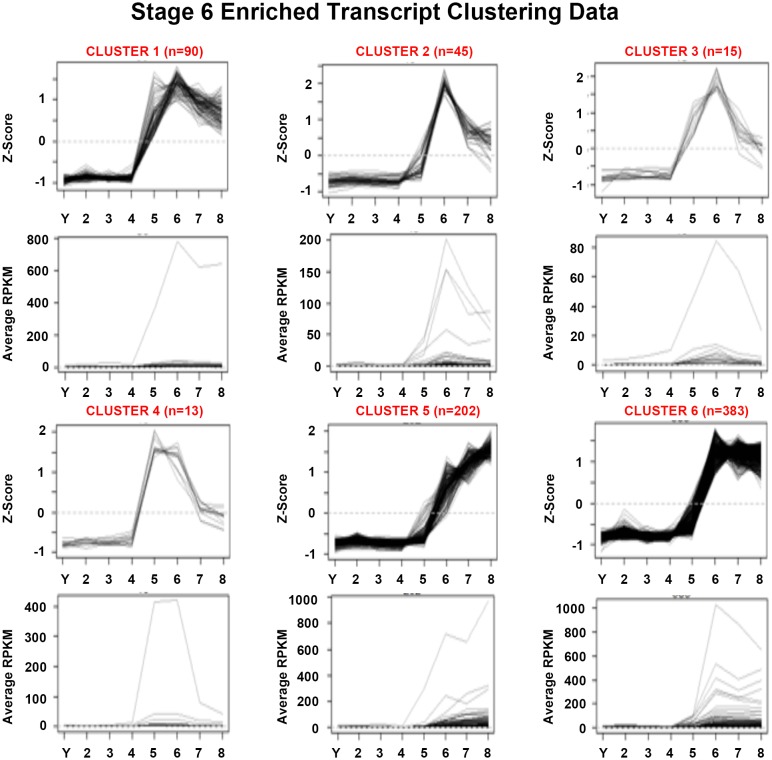
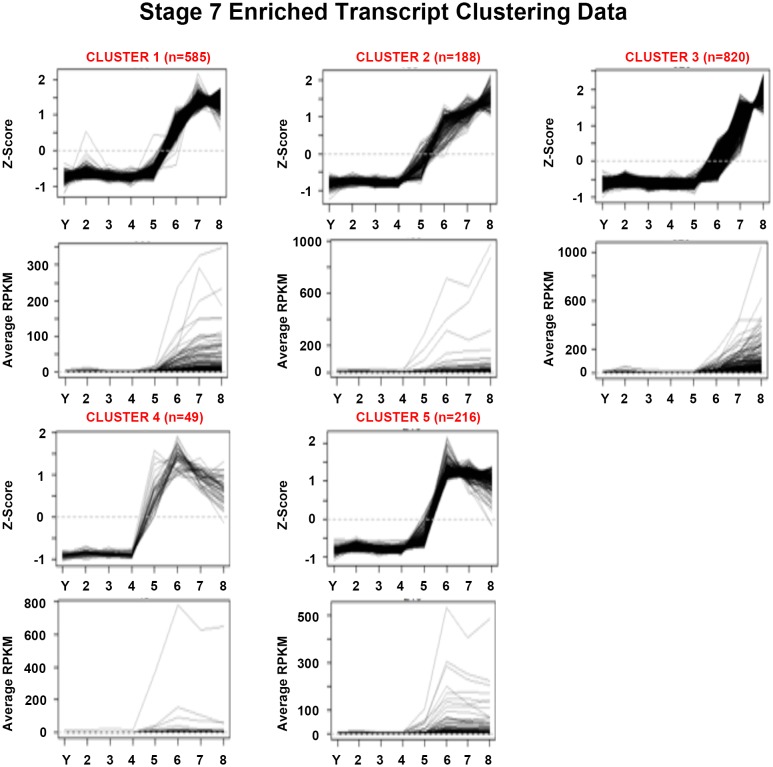
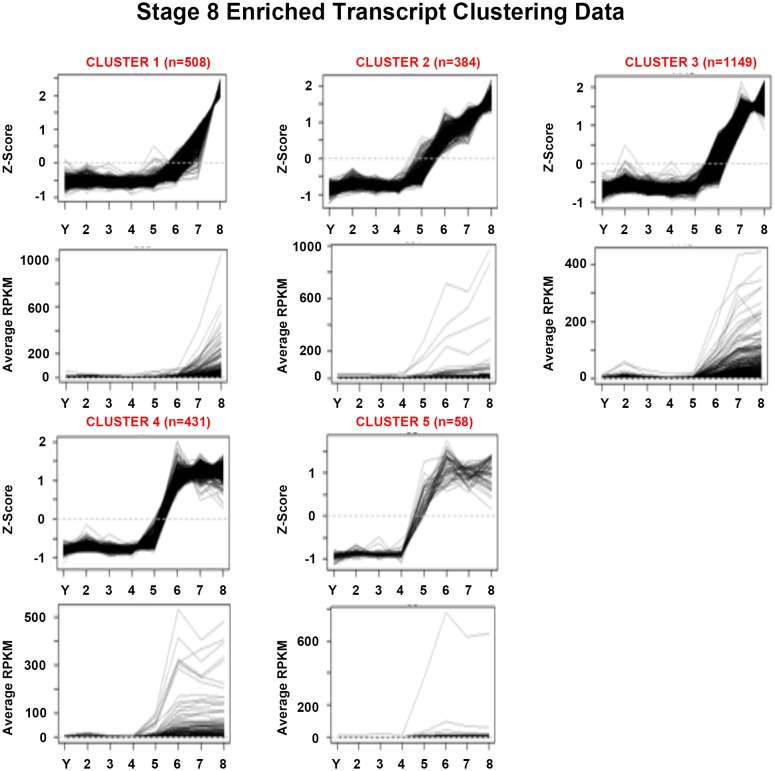
Davies Erin L, Lei Kai, Seidel Christopher W, Kroesen Amanda E, McKinney Sean A, Guo Longhua, Robb Sofia Mc, Ross Eric J, Gotting Kirsten, Alvarado Alejandro Sánchez
Howard Hughes Medical Institute, Stowers Institute for Medical Research, Kansas City, United States.
Elife. 2017 Jan 10;6:e21052. doi: 10.7554/eLife.21052.
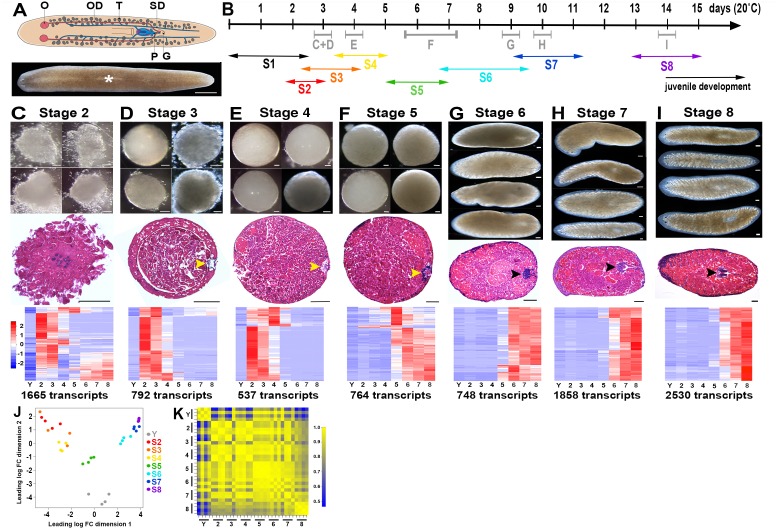
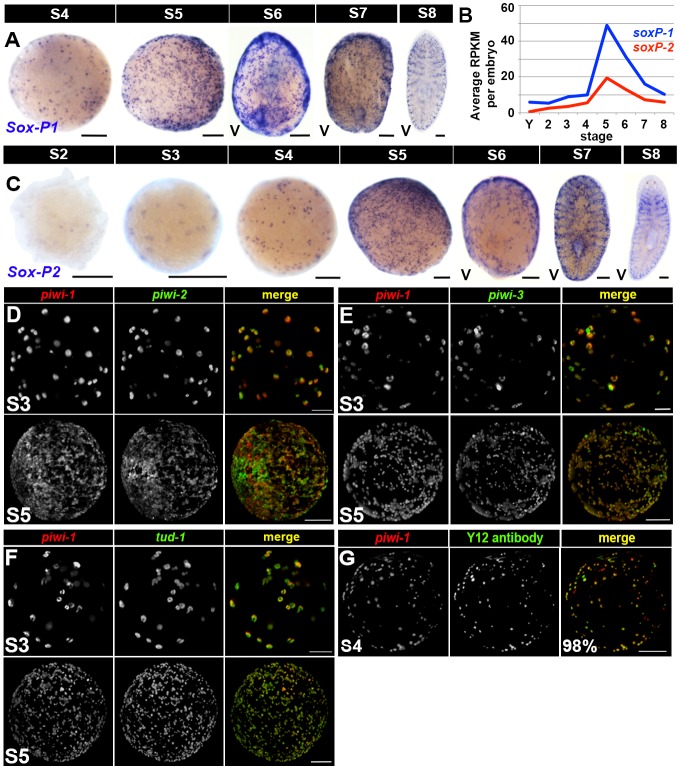
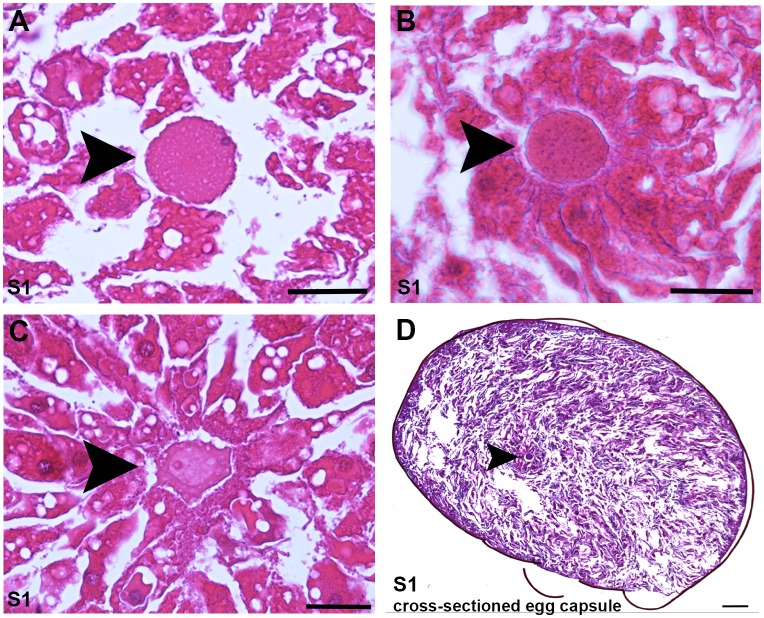
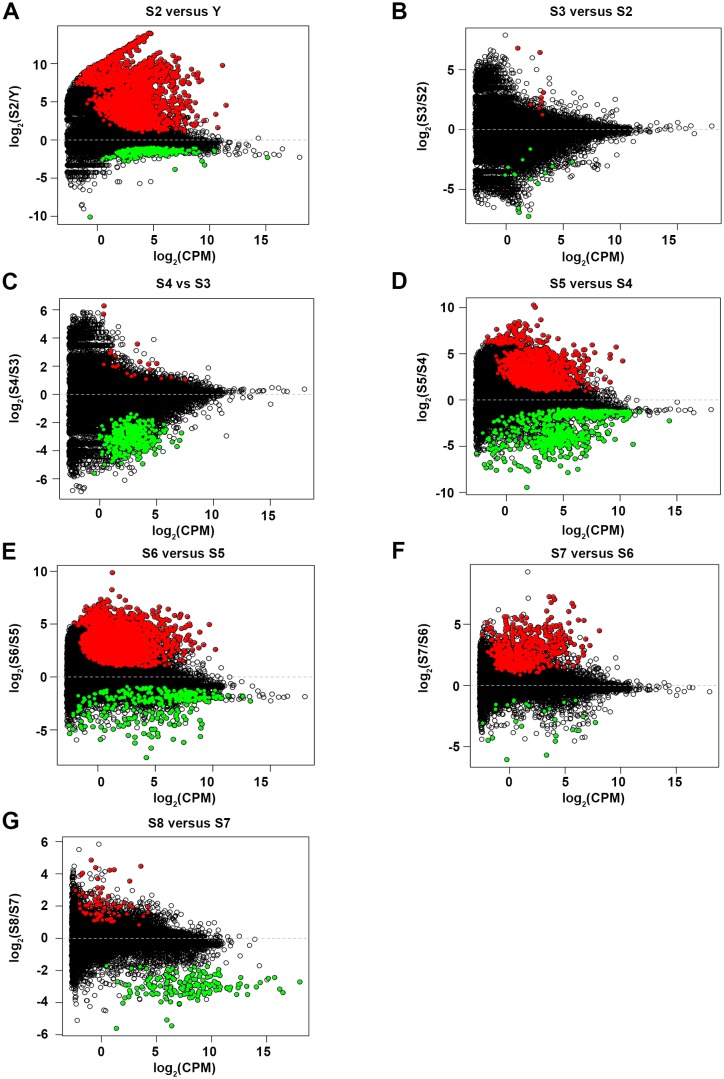
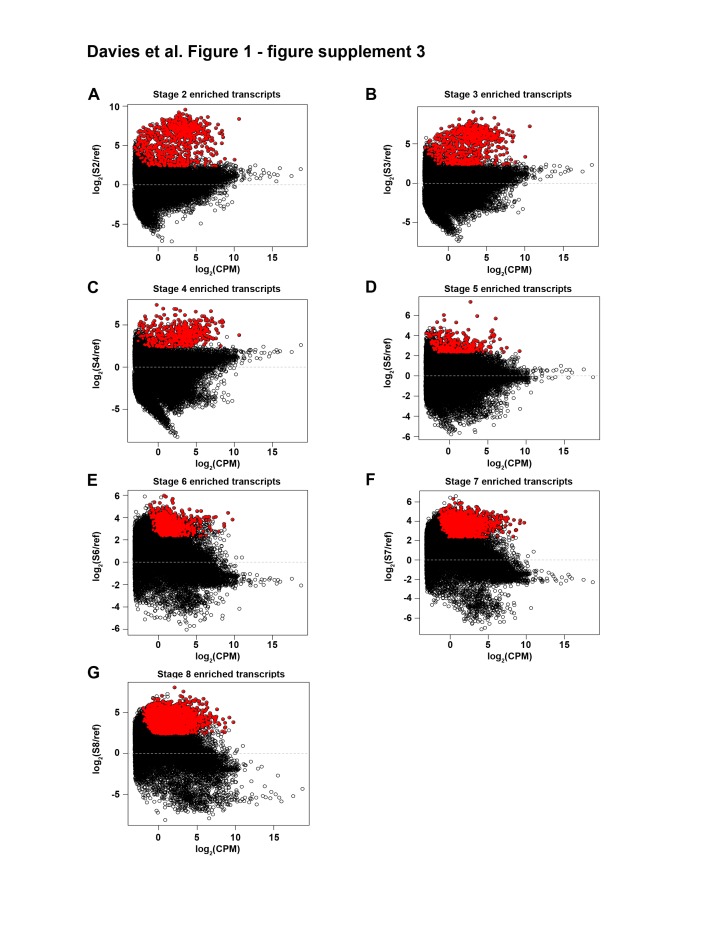
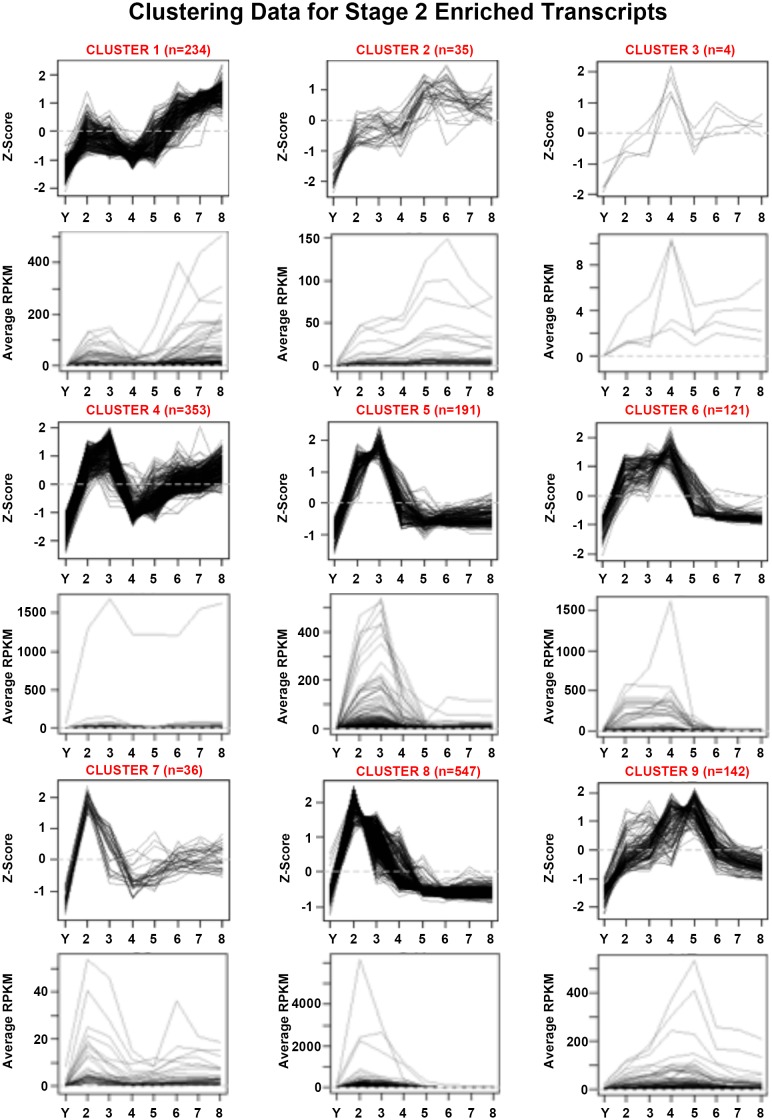
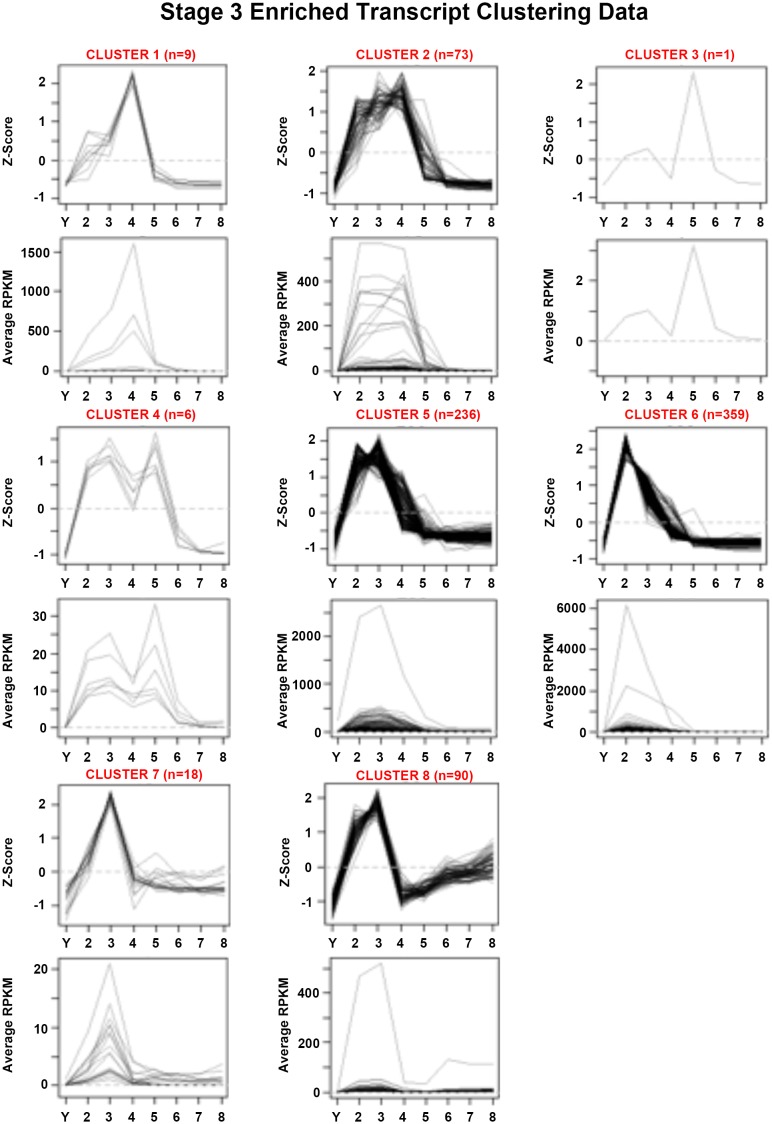
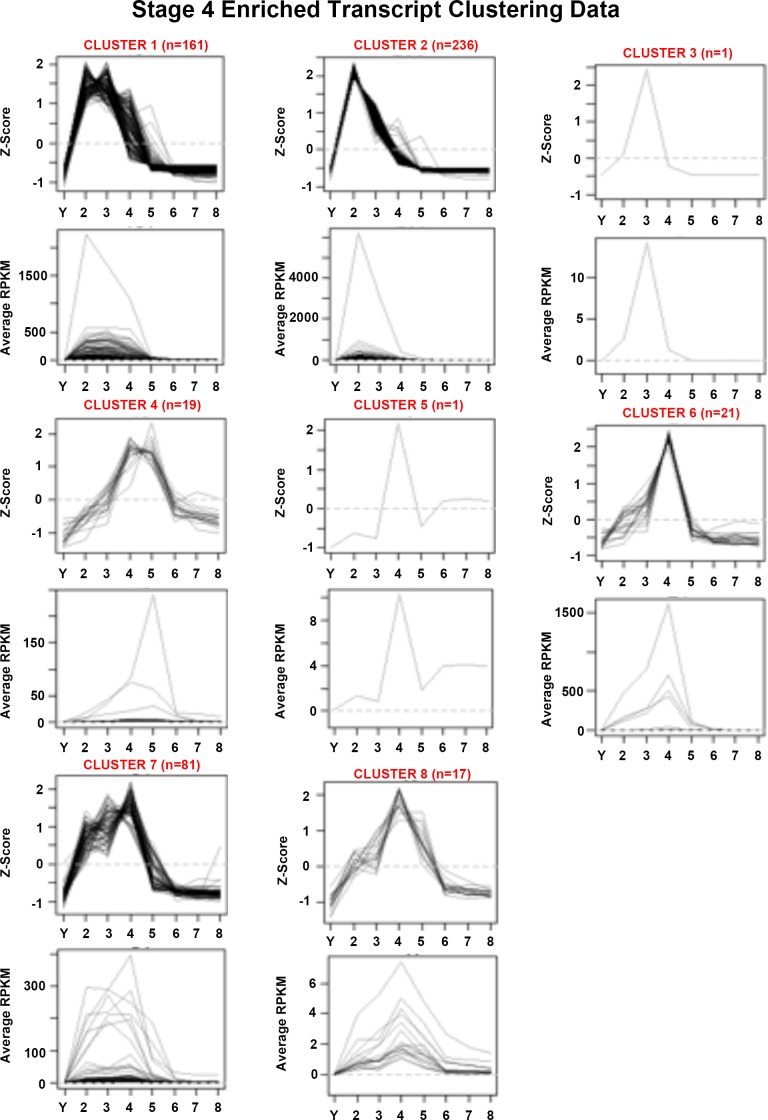
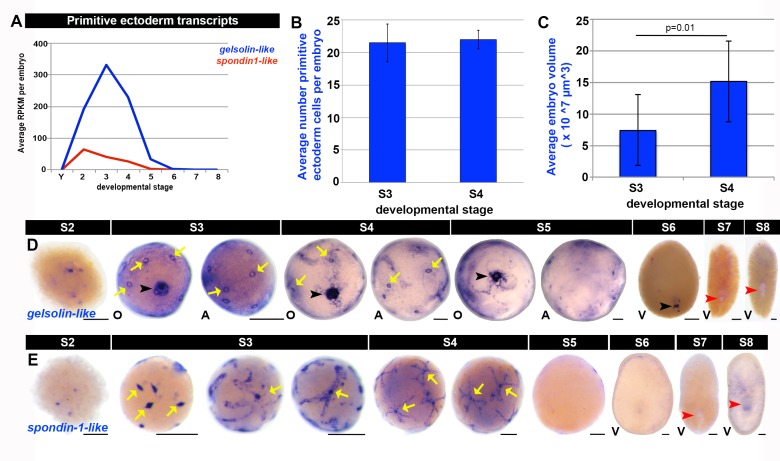
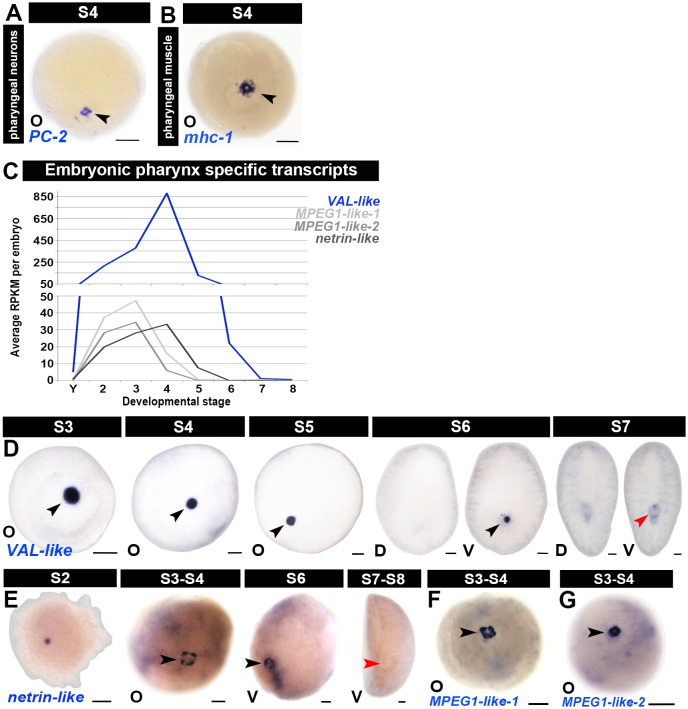
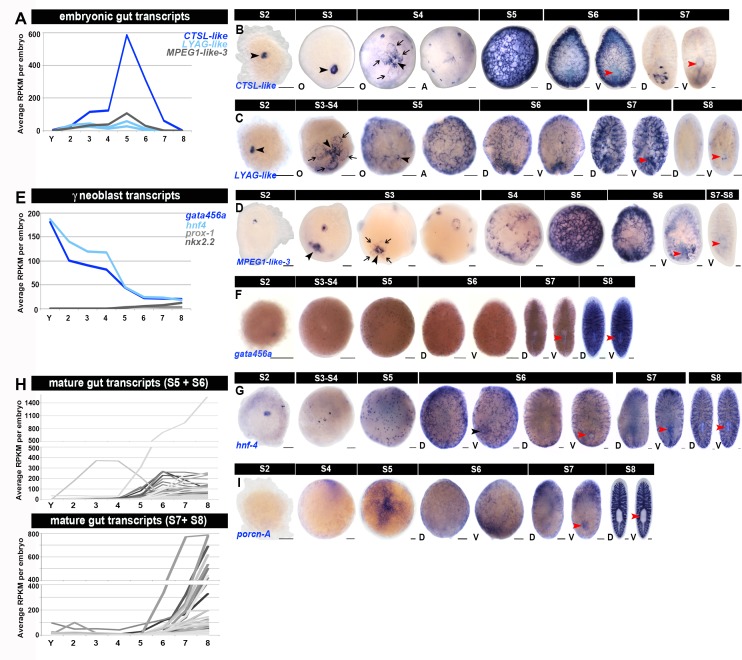
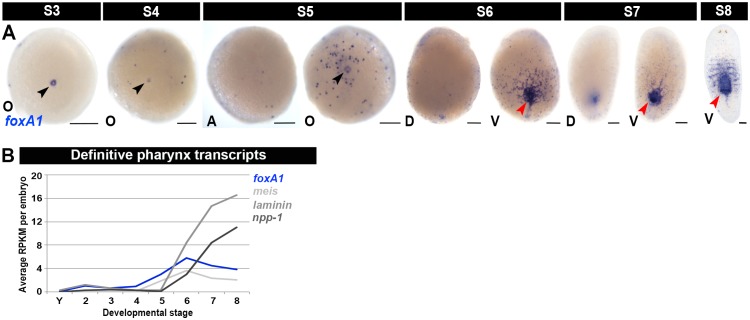
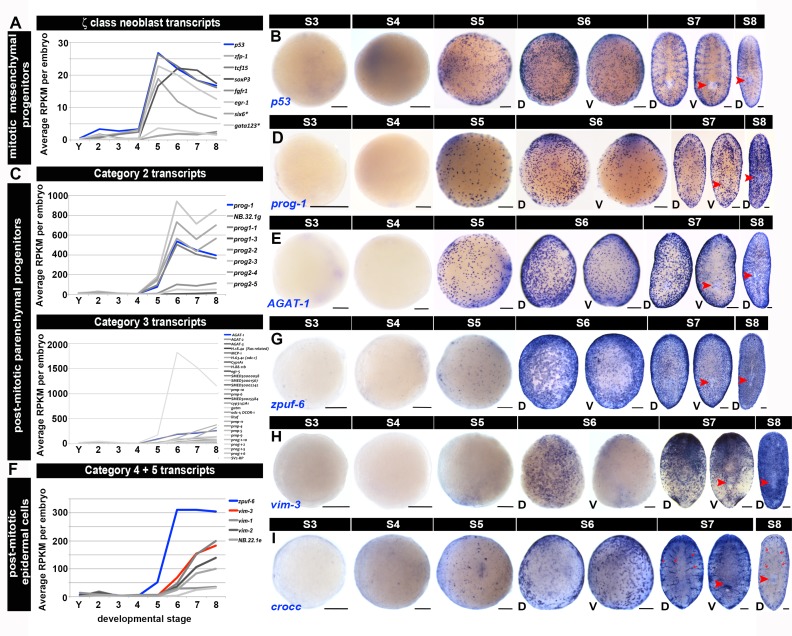
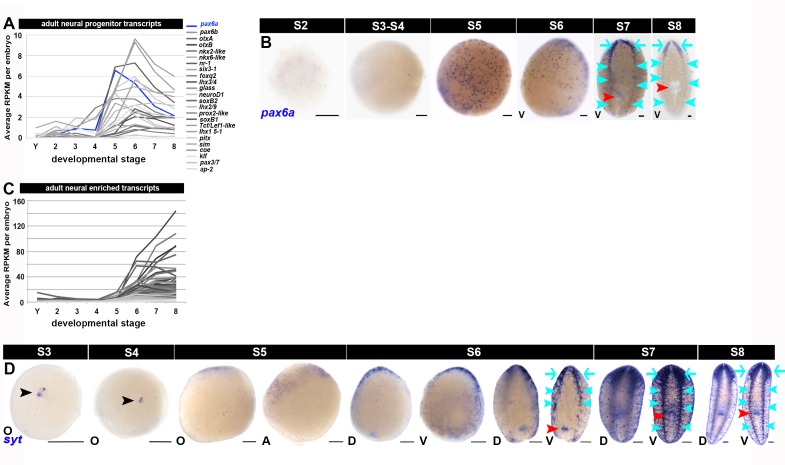
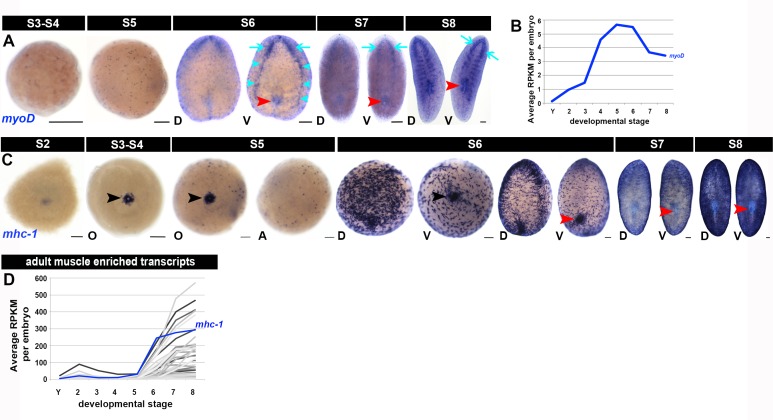
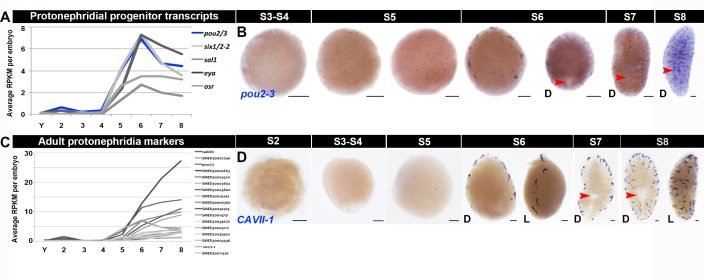
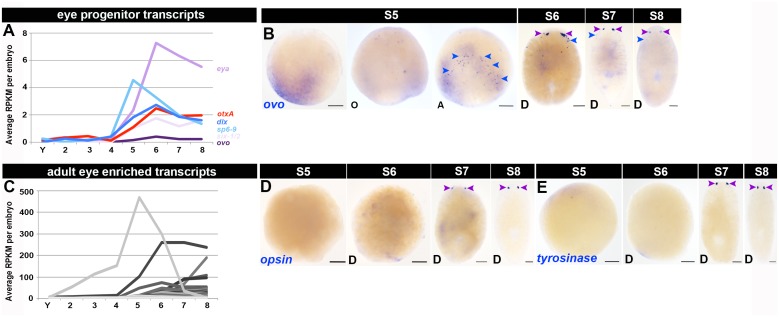
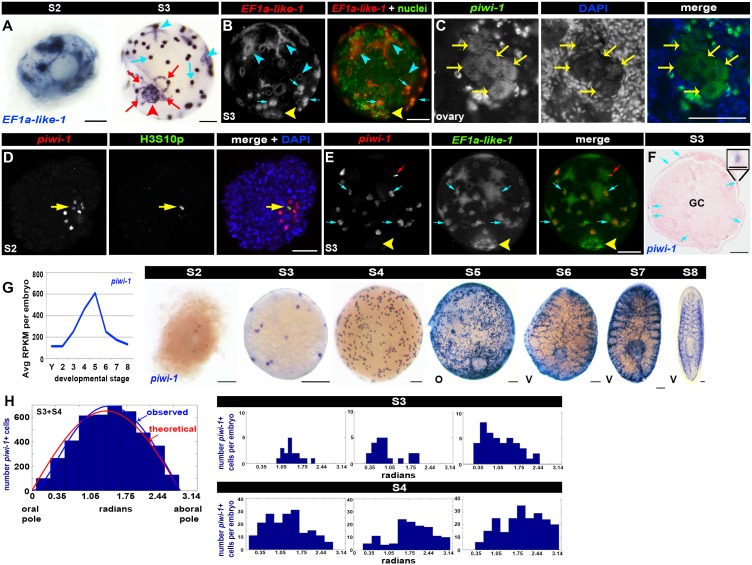
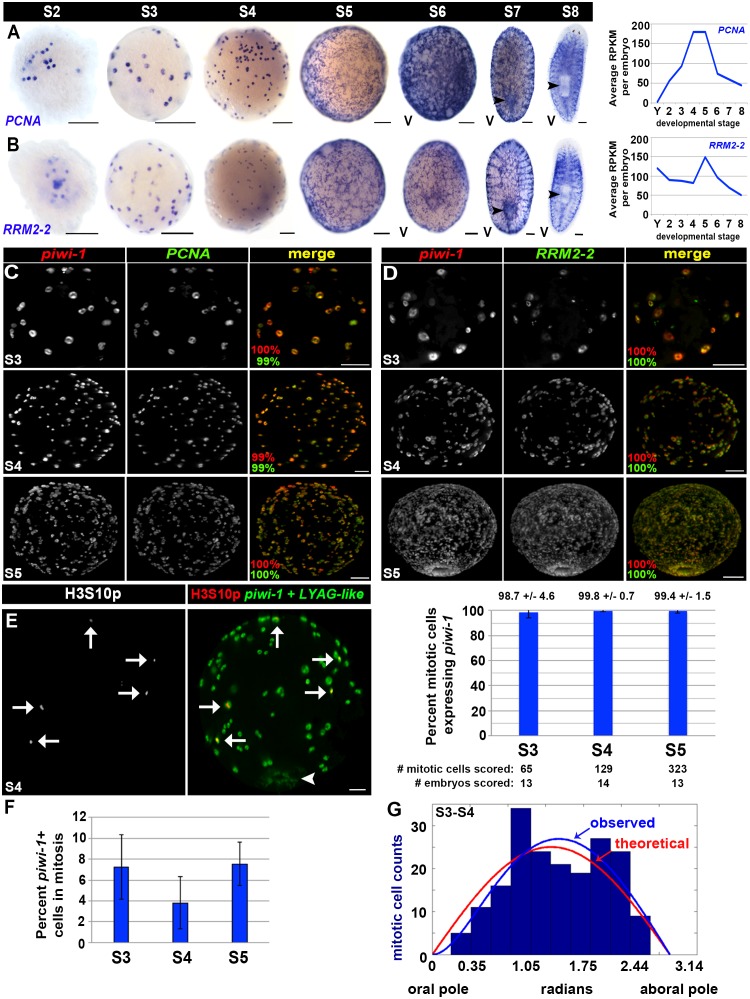
Planarian neoblasts are pluripotent, adult somatic stem cells and lineage-primed progenitors that are required for the production and maintenance of all differentiated cell types, including the germline. Neoblasts, originally defined as undifferentiated cells residing in the adult parenchyma, are frequently compared to embryonic stem cells yet their developmental origin remains obscure. We investigated the provenance of neoblasts during embryogenesis, and report that neoblasts arise from an anarchic, cycling population wholly responsible for production of all temporary and definitive organs during embryogenesis. Early embryonic cells are molecularly and functionally distinct from neoblasts: they express unique cohorts of early embryo enriched transcripts and behave differently than neoblasts in cell transplantation assays. Neoblast lineages arise as organogenesis begins and are required for construction of all major organ systems during embryogenesis. These subpopulations are continuously generated during adulthood, where they act as agents of tissue homeostasis and regeneration.
涡虫新细胞是多能的成体体细胞干细胞和谱系定向祖细胞,是产生和维持包括生殖系在内的所有分化细胞类型所必需的。新细胞最初被定义为存在于成体实质中的未分化细胞,常被与胚胎干细胞相比较,但其发育起源仍不清楚。我们研究了胚胎发生过程中新细胞的来源,并报告新细胞起源于一个无序循环的群体,该群体完全负责胚胎发生过程中所有临时和确定器官的产生。早期胚胎细胞在分子和功能上与新细胞不同:它们表达独特的早期胚胎富集转录本群体,并且在细胞移植试验中的行为与新细胞不同。新细胞谱系在器官发生开始时出现,是胚胎发生过程中构建所有主要器官系统所必需的。这些亚群在成年期持续产生,在那里它们作为组织稳态和再生的因子发挥作用。