Grove Matthew, Kim Hyukmin, Santerre Maryline, Krupka Alexander J, Han Seung Baek, Zhai Jinbin, Cho Jennifer Y, Park Raehee, Harris Michele, Kim Seonhee, Sawaya Bassel E, Kang Shin H, Barbe Mary F, Cho Seo-Hee, Lemay Michel A, Son Young-Jin
Shriners Hospitals Pediatric Research Center, Center for Neural Repair, Lewis Katz School of Medicine, Temple University, Philadelphia, United States.
Department of Anatomy and Cell Biology, Lewis Katz School of Medicine, Temple University, Philadelphia, United States.
Elife. 2017 Jan 26;6:e20982. doi: 10.7554/eLife.20982.
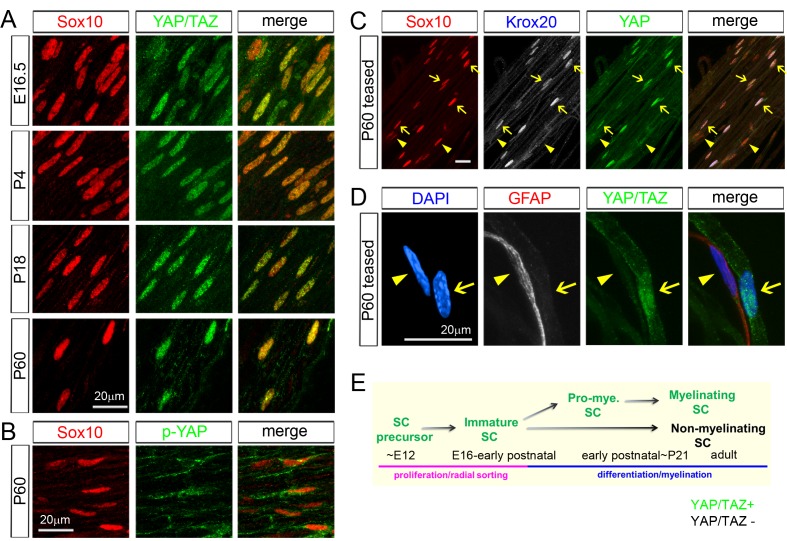
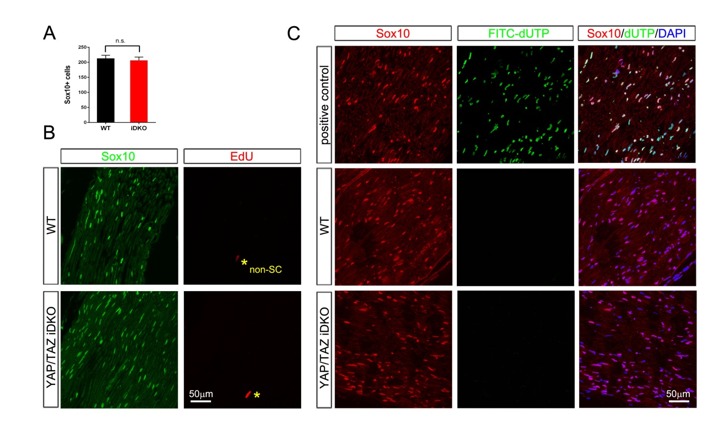
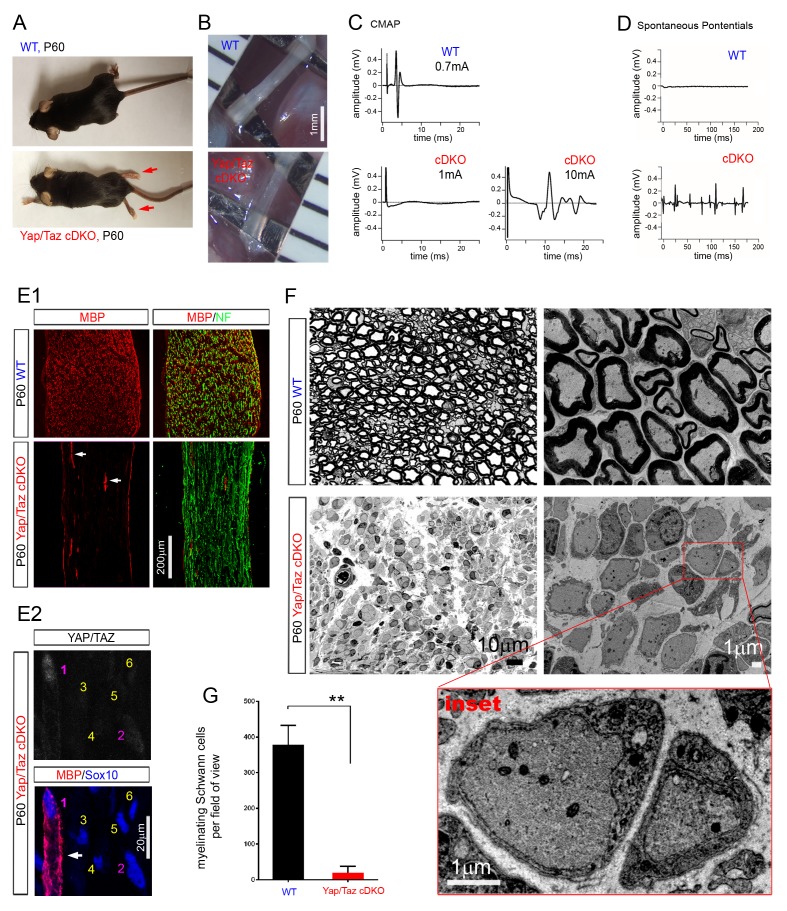
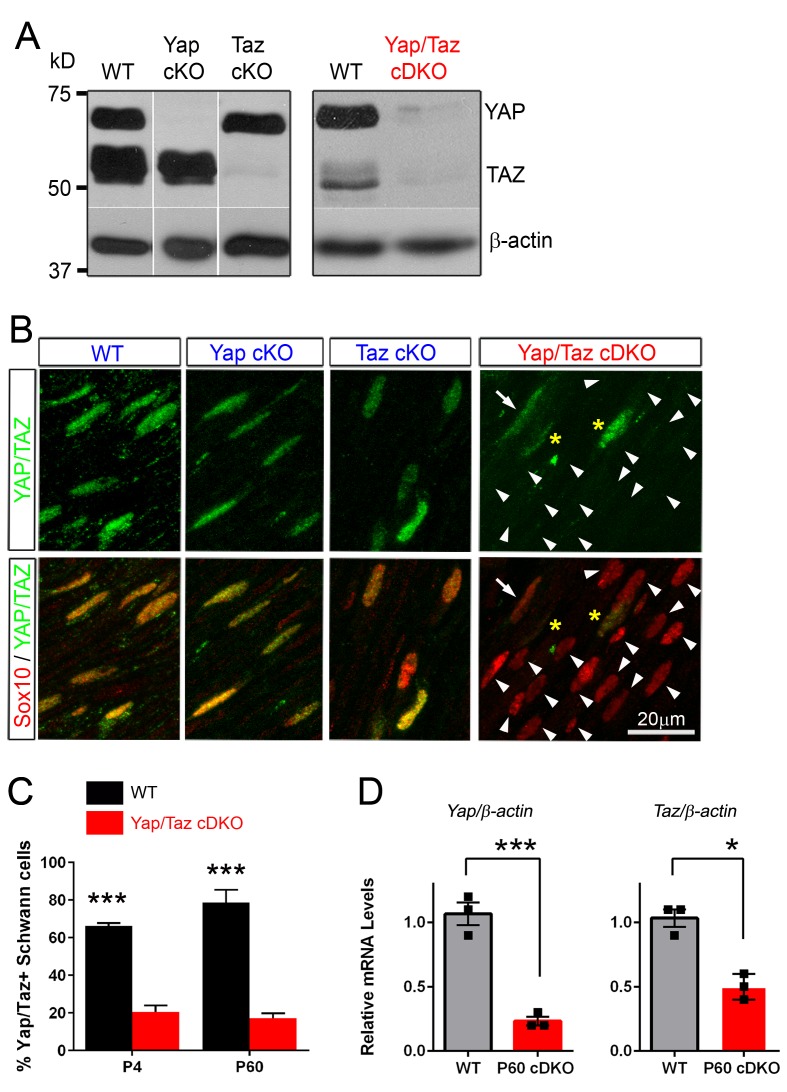
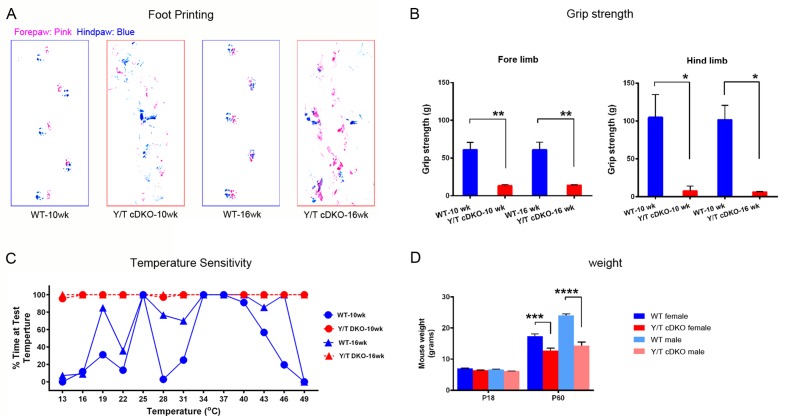
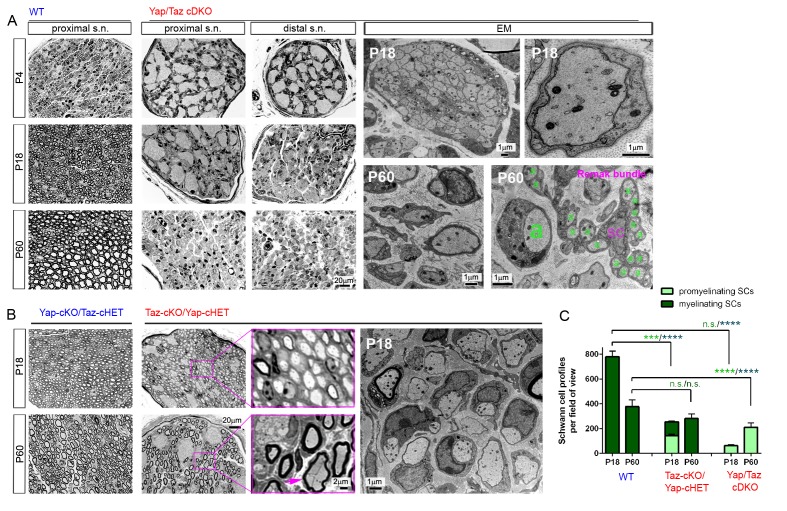
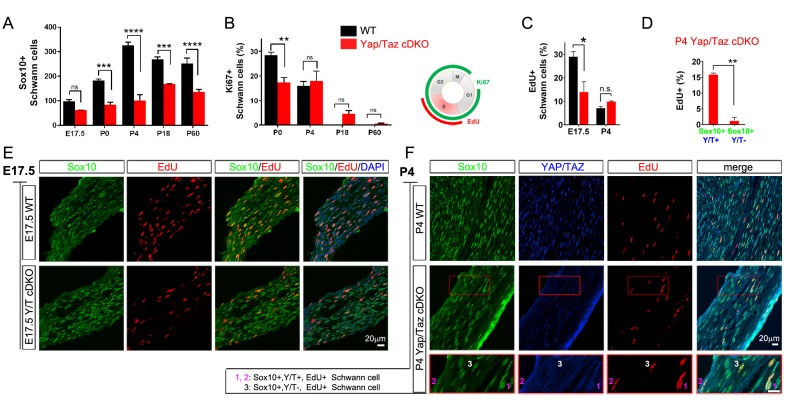
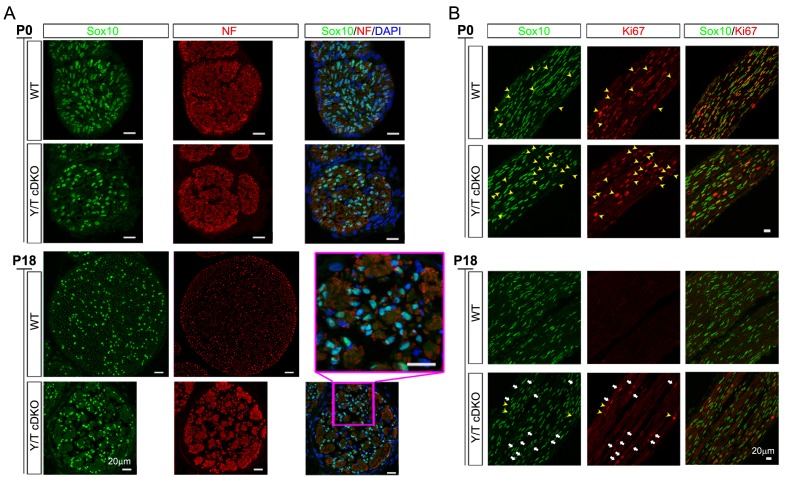
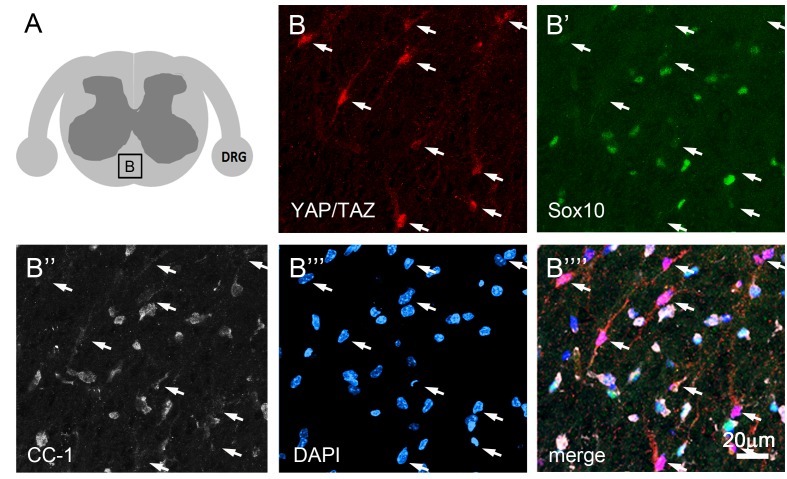
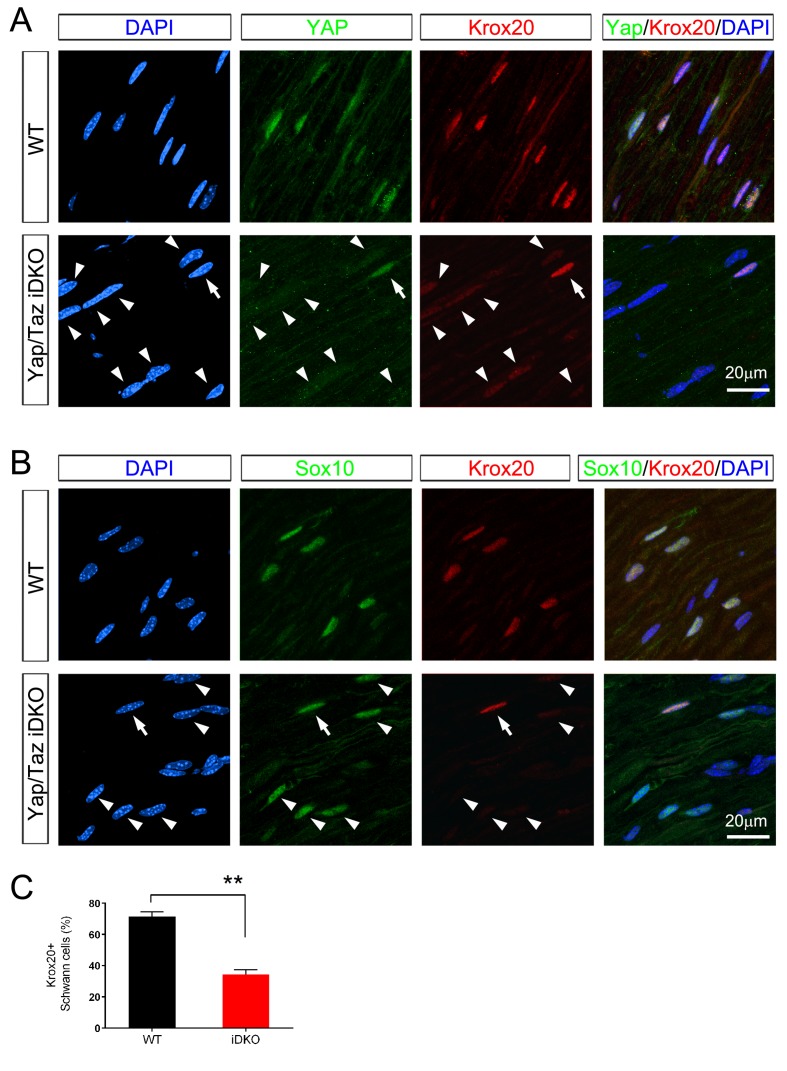
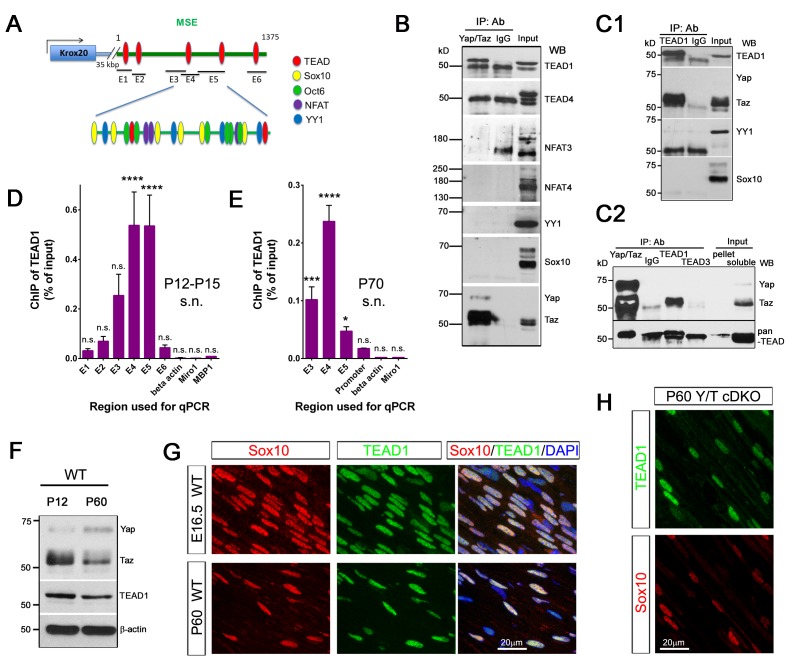
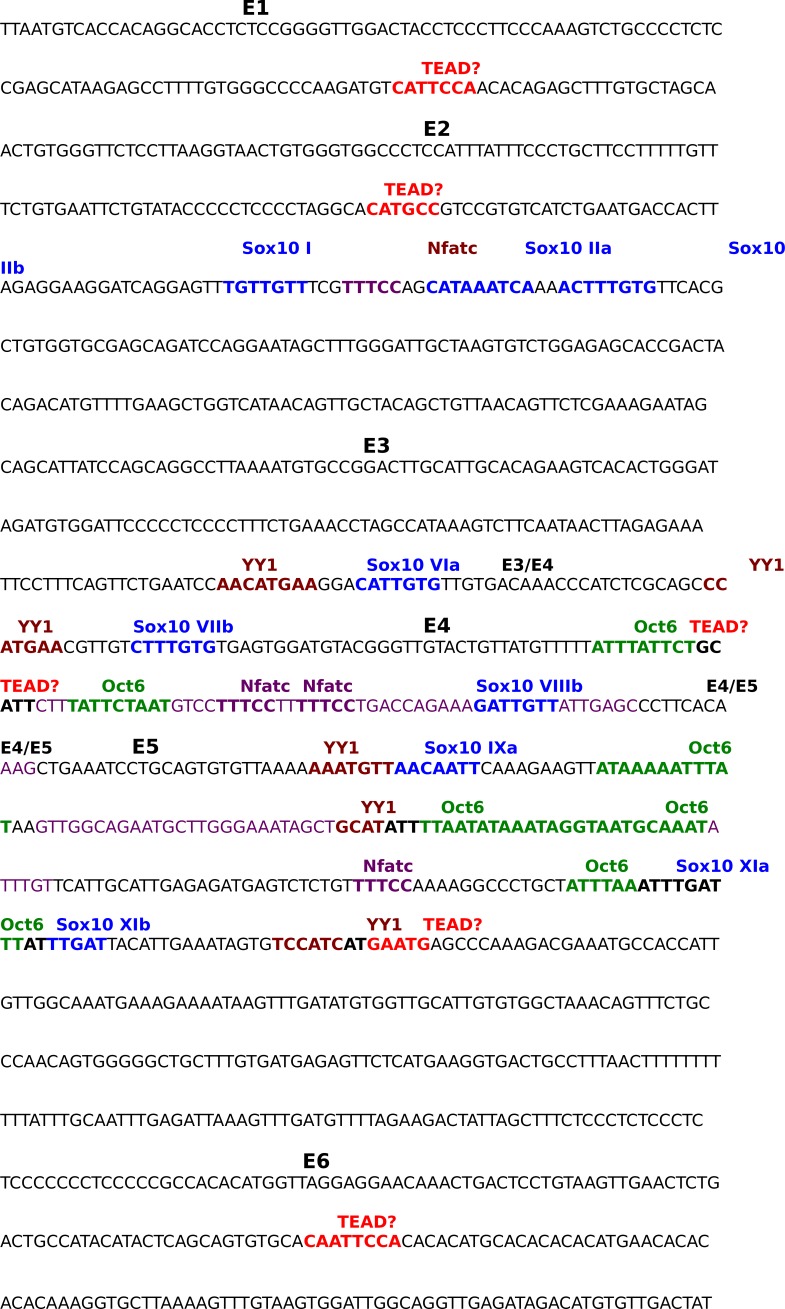
Nuclear exclusion of the transcriptional regulators and potent oncoproteins, YAP/TAZ, is considered necessary for adult tissue homeostasis. Here we show that nuclear YAP/TAZ are essential regulators of peripheral nerve development and myelin maintenance. To proliferate, developing Schwann cells (SCs) require YAP/TAZ to enter S-phase and, without them, fail to generate sufficient SCs for timely axon sorting. To differentiate, SCs require YAP/TAZ to upregulate Krox20 and, without them, completely fail to myelinate, resulting in severe peripheral neuropathy. Remarkably, in adulthood, nuclear YAP/TAZ are selectively expressed by myelinating SCs, and conditional ablation results in severe peripheral demyelination and mouse death. YAP/TAZ regulate both developmental and adult myelination by driving TEAD1 to activate Krox20. Therefore, YAP/TAZ are crucial for SCs to myelinate developing nerve and to maintain myelinated nerve in adulthood. Our study also provides a new insight into the role of nuclear YAP/TAZ in homeostatic maintenance of an adult tissue.
转录调节因子及强效癌蛋白YAP/TAZ的核排除被认为是成年组织稳态所必需的。在此我们表明,核内YAP/TAZ是外周神经发育和髓鞘维持的关键调节因子。为了增殖,发育中的雪旺细胞(SCs)需要YAP/TAZ进入S期,没有它们,就无法产生足够的雪旺细胞来及时进行轴突分选。为了分化,雪旺细胞需要YAP/TAZ上调Krox20,没有它们,就会完全无法形成髓鞘,导致严重的外周神经病变。值得注意的是,在成年期,核内YAP/TAZ由形成髓鞘的雪旺细胞选择性表达,条件性消融会导致严重的外周脱髓鞘和小鼠死亡。YAP/TAZ通过驱动TEAD1激活Krox20来调节发育和成年期的髓鞘形成。因此,YAP/TAZ对于雪旺细胞形成发育中的神经髓鞘以及在成年期维持有髓神经至关重要。我们的研究还为核内YAP/TAZ在成年组织稳态维持中的作用提供了新的见解。