Maiato Helder, Gomes Ana Margarida, Sousa Filipe, Barisic Marin
Chromosome Instability & Dynamics Laboratory, Instituto de Biologia Molecular e Celular, Universidade do Porto, Rua Alfredo Allen 208, 4200-135 Porto, Portugal.
Instituto de Investigação e Inovação em Saúde-i3S, Universidade do Porto, Rua Alfredo Allen 208, 4200-135 Porto, Portugal.
Biology (Basel). 2017 Feb 17;6(1):13. doi: 10.3390/biology6010013.
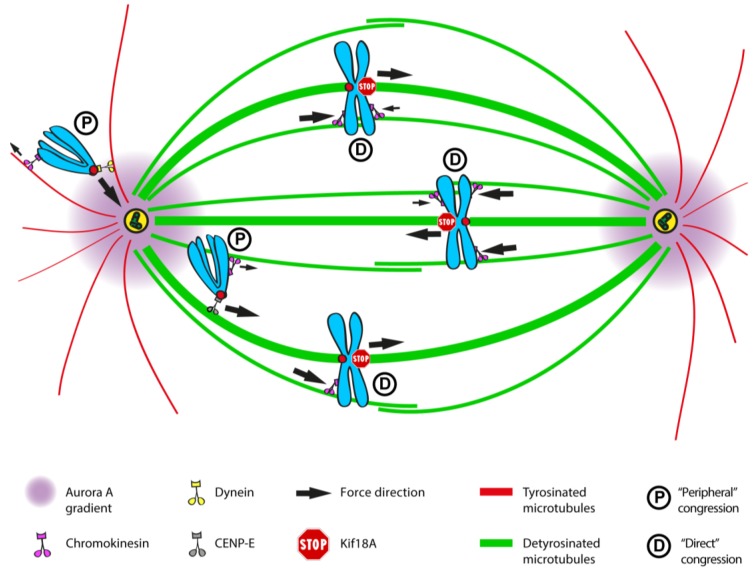
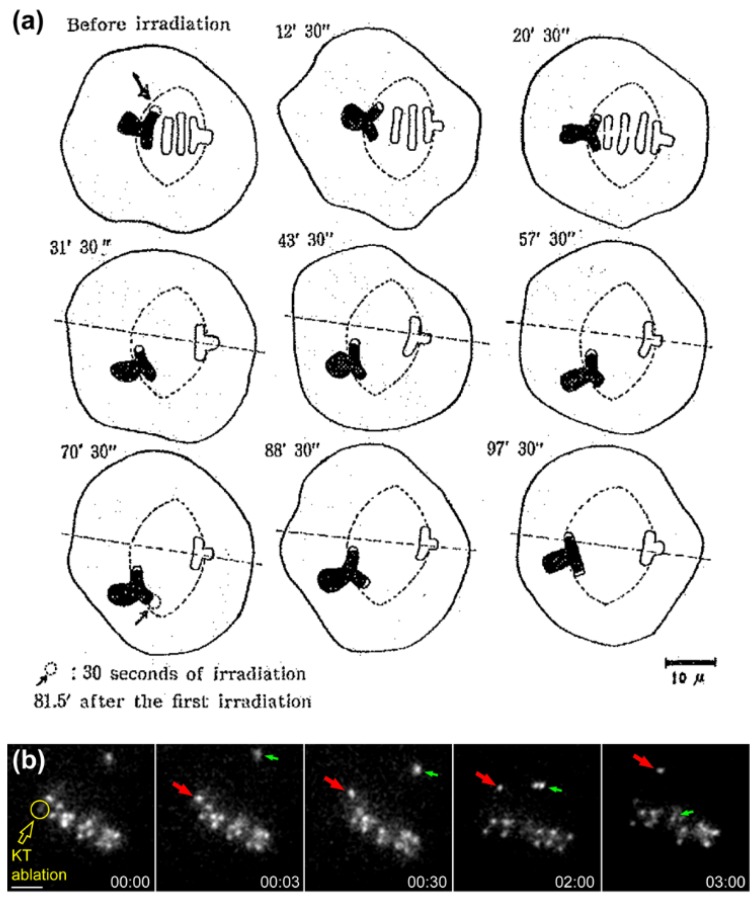
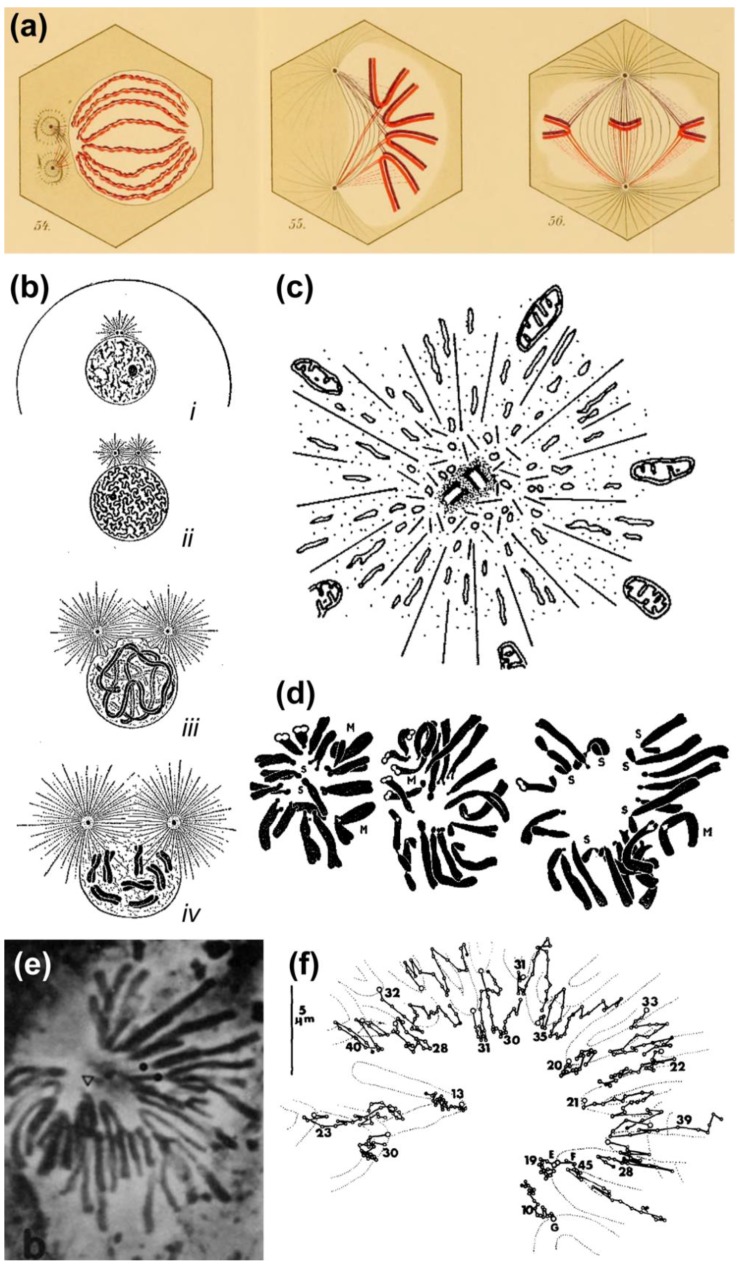
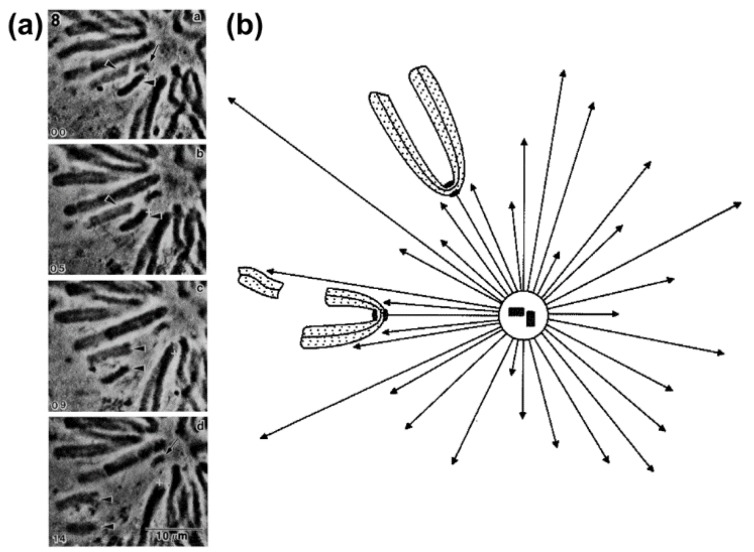
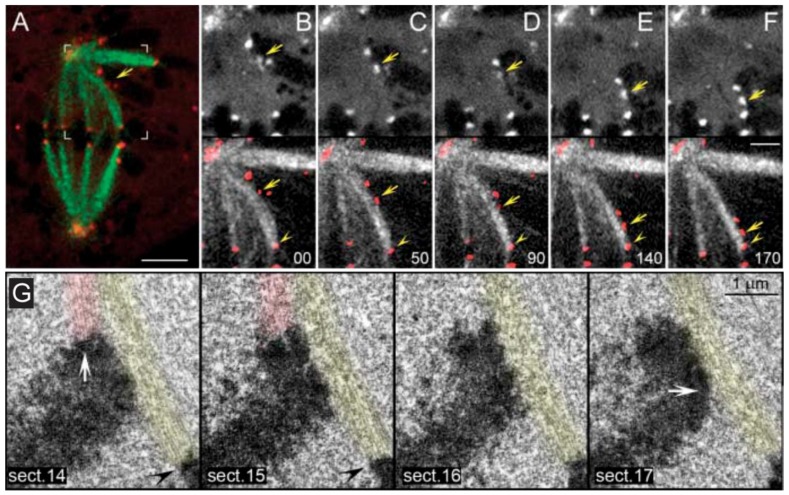
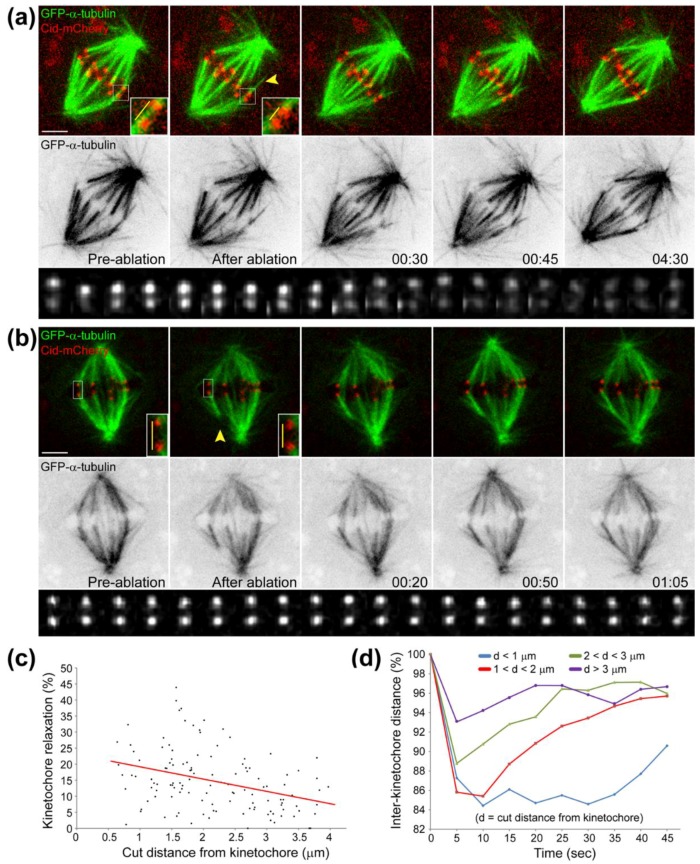
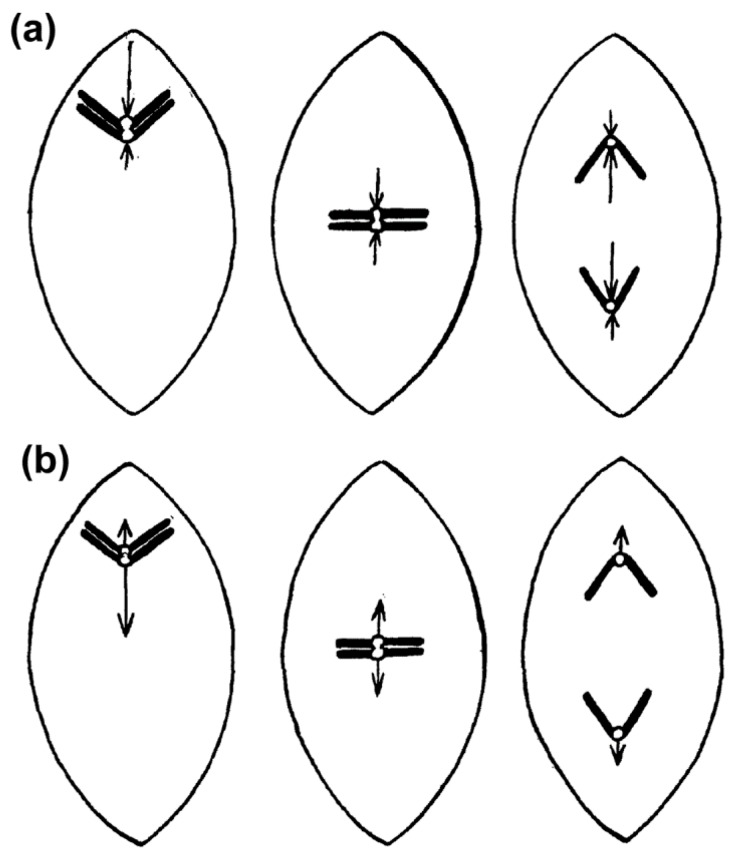
Chromosome congression during prometaphase culminates with the establishment of a metaphase plate, a hallmark of mitosis in metazoans. Classical views resulting from more than 100 years of research on this topic have attempted to explain chromosome congression based on the balance between opposing pulling and/or pushing forces that reach an equilibrium near the spindle equator. However, in mammalian cells, chromosome bi-orientation and force balance at kinetochores are not required for chromosome congression, whereas the mechanisms of chromosome congression are not necessarily involved in the maintenance of chromosome alignment after congression. Thus, chromosome congression and maintenance of alignment are determined by different principles. Moreover, it is now clear that not all chromosomes use the same mechanism for congressing to the spindle equator. Those chromosomes that are favorably positioned between both poles when the nuclear envelope breaks down use the so-called "direct congression" pathway in which chromosomes align after bi-orientation and the establishment of end-on kinetochore-microtubule attachments. This favors the balanced action of kinetochore pulling forces and polar ejection forces along chromosome arms that drive chromosome oscillatory movements during and after congression. The other pathway, which we call "peripheral congression", is independent of end-on kinetochore microtubule-attachments and relies on the dominant and coordinated action of the kinetochore motors Dynein and Centromere Protein E (CENP-E) that mediate the lateral transport of peripheral chromosomes along microtubules, first towards the poles and subsequently towards the equator. How the opposite polarities of kinetochore motors are regulated in space and time to drive congression of peripheral chromosomes only now starts to be understood. This appears to be regulated by position-dependent phosphorylation of both Dynein and CENP-E and by spindle microtubule diversity by means of tubulin post-translational modifications. This so-called "tubulin code" might work as a navigation system that selectively guides kinetochore motors with opposite polarities along specific spindle microtubule populations, ultimately leading to the congression of peripheral chromosomes. We propose an integrated model of chromosome congression in mammalian cells that depends essentially on the following parameters: (1) chromosome position relative to the spindle poles after nuclear envelope breakdown; (2) establishment of stable end-on kinetochore-microtubule attachments and bi-orientation; (3) coordination between kinetochore- and arm-associated motors; and (4) spatial signatures associated with post-translational modifications of specific spindle microtubule populations. The physiological consequences of abnormal chromosome congression, as well as the therapeutic potential of inhibiting chromosome congression are also discussed.
前中期的染色体汇聚以中期板的形成为顶点,中期板是后生动物有丝分裂的一个标志。基于对该主题100多年的研究得出的传统观点试图根据在纺锤体赤道附近达到平衡的相反拉力和/或推力之间的平衡来解释染色体汇聚。然而,在哺乳动物细胞中,染色体向两极定向和动粒处的力平衡对于染色体汇聚并非必需,而染色体汇聚的机制不一定参与汇聚后染色体排列的维持。因此,染色体汇聚和排列的维持由不同的原理决定。此外,现在很清楚,并非所有染色体都使用相同的机制汇聚到纺锤体赤道。那些在核膜破裂时位于两极之间有利位置的染色体使用所谓的“直接汇聚”途径,即染色体在向两极定向和端对端动粒 - 微管附着建立后排列。这有利于动粒拉力和沿着染色体臂的极性弹射力的平衡作用,这些力在汇聚期间和之后驱动染色体的振荡运动。另一条途径,我们称之为“周边汇聚”,独立于端对端动粒微管附着,并且依赖于动粒马达动力蛋白和着丝粒蛋白E(CENP - E)的主导和协调作用,它们介导周边染色体沿着微管的侧向运输,首先朝向两极,随后朝向赤道。动粒马达相反极性如何在空间和时间上受到调节以驱动周边染色体的汇聚,直到现在才开始被理解。这似乎通过动力蛋白和CENP - E的位置依赖性磷酸化以及通过微管蛋白翻译后修饰导致的纺锤体微管多样性来调节。这种所谓的“微管蛋白密码”可能作为一种导航系统,选择性地引导具有相反极性的动粒马达沿着特定的纺锤体微管群体移动,最终导致周边染色体的汇聚。我们提出了一个哺乳动物细胞中染色体汇聚的综合模型,该模型主要依赖于以下参数:(1)核膜破裂后染色体相对于纺锤体两极的位置;(2)稳定的端对端动粒 - 微管附着和向两极定向的建立;(3)动粒和臂相关马达之间的协调;以及(4)与特定纺锤体微管群体翻译后修饰相关的空间特征。还讨论了异常染色体汇聚的生理后果以及抑制染色体汇聚的治疗潜力。