Neurosciences and Mental Health, The Hospital for Sick Children, Toronto, Canada.
Department of Physiology and the Institute of Biomaterials and Biomedical Engineering, University of Toronto, Toronto, Canada.
J Physiol. 2018 May 1;596(9):1681-1697. doi: 10.1113/JP275240. Epub 2018 Feb 21.
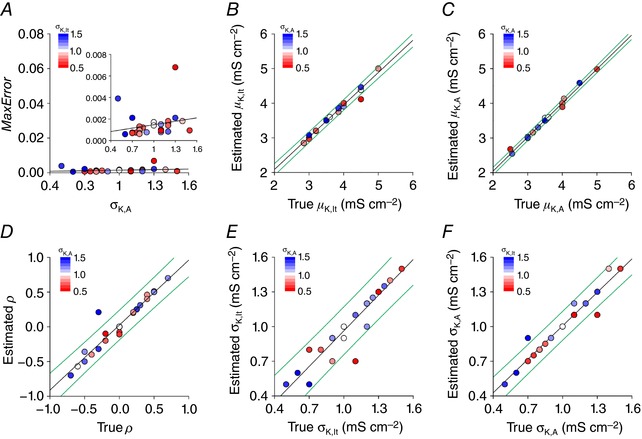
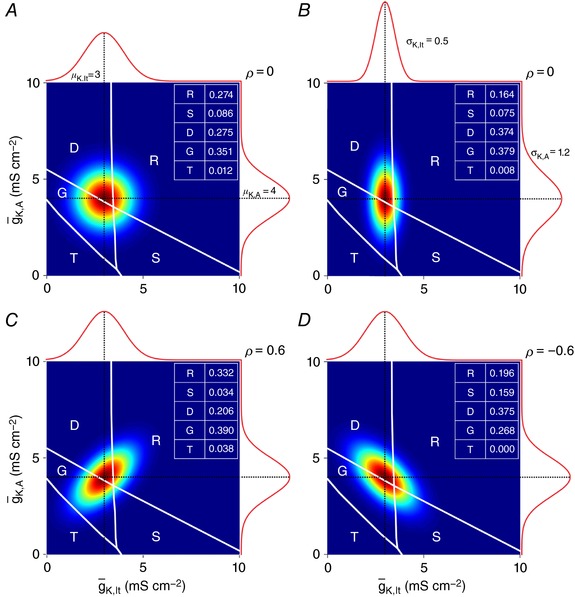
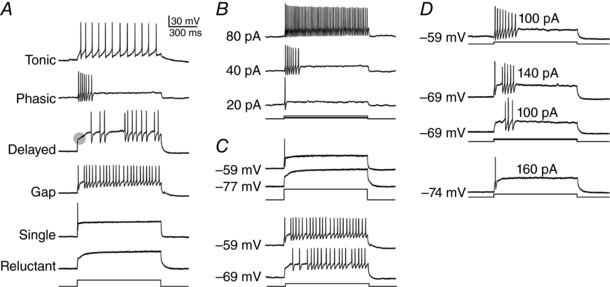
Distinct spiking patterns may arise from qualitative differences in ion channel expression (i.e. when different neurons express distinct ion channels) and/or when quantitative differences in expression levels qualitatively alter the spike generation process. We hypothesized that spiking patterns in neurons of the superficial dorsal horn (SDH) of spinal cord reflect both mechanisms. We reproduced SDH neuron spiking patterns by varying densities of K 1- and A-type potassium conductances. Plotting the spiking patterns that emerge from different density combinations revealed spiking-pattern regions separated by boundaries (bifurcations). This map suggests that certain spiking pattern combinations occur when the distribution of potassium channel densities straddle boundaries, whereas other spiking patterns reflect distinct patterns of ion channel expression. The former mechanism may explain why certain spiking patterns co-occur in genetically identified neuron types. We also present algorithms to predict spiking pattern proportions from ion channel density distributions, and vice versa.
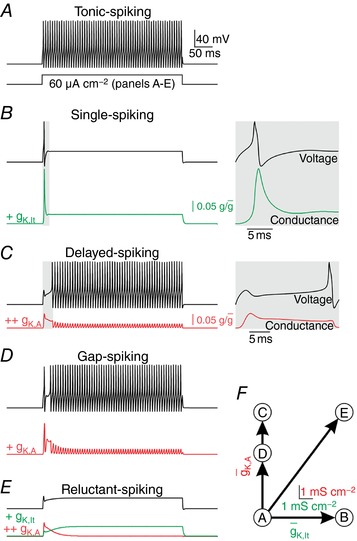
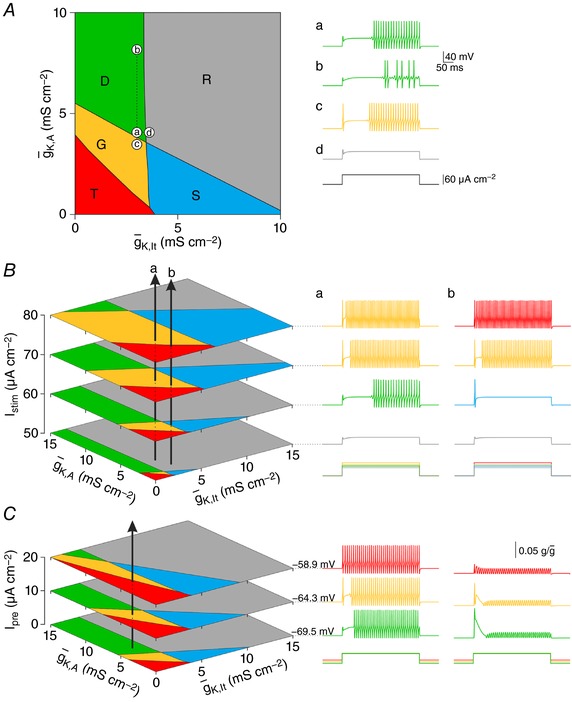
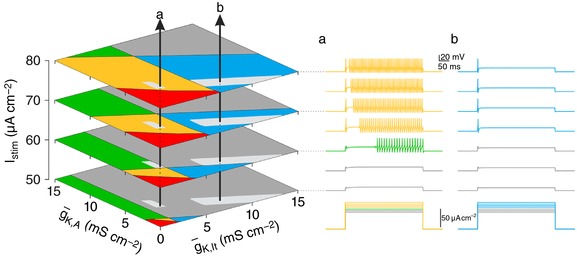
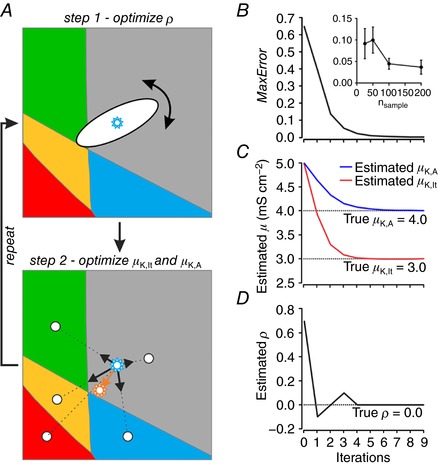
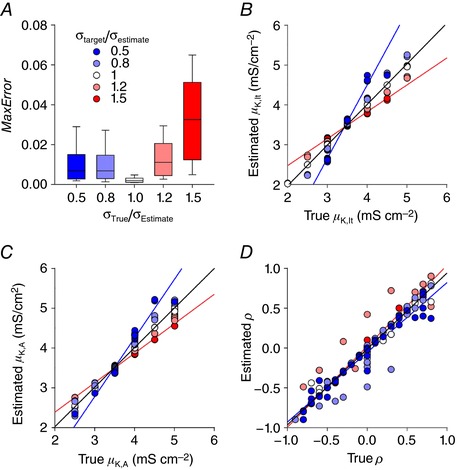
Neurons are often classified by spiking pattern. Yet, some neurons exhibit distinct patterns under subtly different test conditions, which suggests that they operate near an abrupt transition, or bifurcation. A set of such neurons may exhibit heterogeneous spiking patterns not because of qualitative differences in which ion channels they express, but rather because quantitative differences in expression levels cause neurons to operate on opposite sides of a bifurcation. Neurons in the spinal dorsal horn, for example, respond to somatic current injection with patterns that include tonic, single, gap, delayed and reluctant spiking. It is unclear whether these patterns reflect five cell populations (defined by distinct ion channel expression patterns), heterogeneity within a single population, or some combination thereof. We reproduced all five spiking patterns in a computational model by varying the densities of a low-threshold (K 1-type) potassium conductance and an inactivating (A-type) potassium conductance and found that single, gap, delayed and reluctant spiking arise when the joint probability distribution of those channel densities spans two intersecting bifurcations that divide the parameter space into quadrants, each associated with a different spiking pattern. Tonic spiking likely arises from a separate distribution of potassium channel densities. These results argue in favour of two cell populations, one characterized by tonic spiking and the other by heterogeneous spiking patterns. We present algorithms to predict spiking pattern proportions based on ion channel density distributions and, conversely, to estimate ion channel density distributions based on spiking pattern proportions. The implications for classifying cells based on spiking pattern are discussed.
尖峰模式的差异可能源于离子通道表达的定性差异(即不同神经元表达不同的离子通道)和/或表达水平的定量差异定性地改变了尖峰产生过程。我们假设脊髓背角浅层(SDH)神经元的尖峰模式反映了这两种机制。我们通过改变 K 1-和 A 型钾电导的密度来再现 SDH 神经元的尖峰模式。绘制不同密度组合产生的尖峰模式图揭示了由边界(分叉)分隔的尖峰模式区域。该图谱表明,当钾通道密度分布跨越边界时,某些尖峰模式组合会出现,而其他尖峰模式则反映了离子通道表达的不同模式。前一种机制可以解释为什么某些尖峰模式在遗传鉴定的神经元类型中同时出现。我们还提出了从离子通道密度分布预测尖峰模式比例的算法,反之亦然。
神经元通常根据尖峰模式进行分类。然而,一些神经元在略微不同的测试条件下表现出明显不同的模式,这表明它们处于突然转变或分叉的边缘。一组这样的神经元可能表现出异质的尖峰模式,不是因为它们表达的离子通道存在定性差异,而是因为表达水平的定量差异导致神经元在分叉的相反两侧运行。例如,脊髓背角中的神经元对体电流注射的反应模式包括紧张性、单峰、间隙、延迟和勉强性尖峰。这些模式是否反映了五个细胞群体(由不同的离子通道表达模式定义)、单个群体内的异质性,还是它们的某种组合尚不清楚。我们通过改变低阈值(K 1 型)钾电导和失活(A 型)钾电导的密度,在计算模型中再现了所有五种尖峰模式,发现当这些通道密度的联合概率分布跨越两个相交的分叉时,会出现单峰、间隙、延迟和勉强性尖峰,这些分叉将参数空间划分为四个象限,每个象限与不同的尖峰模式相关联。紧张性尖峰可能来自钾通道密度的单独分布。这些结果支持两种细胞群体,一种以紧张性尖峰为特征,另一种以异质尖峰模式为特征。我们提出了基于离子通道密度分布预测尖峰模式比例的算法,反之亦然,基于尖峰模式比例估计离子通道密度分布。讨论了基于尖峰模式对细胞进行分类的意义。