Oxford Centre for Diabetes, Endocrinology and Metabolism, Radcliffe Department of Medicine, University of Oxford, Churchill Hospital, Oxford, OX3 7LE, UK.
Metabolic Physiology, Department of Neuroscience and Physiology, Medicinaregatan 11, Gothenburg, S-413 09, Sweden.
J Physiol. 2018 May 1;596(9):1601-1626. doi: 10.1113/JP275587. Epub 2018 Mar 30.
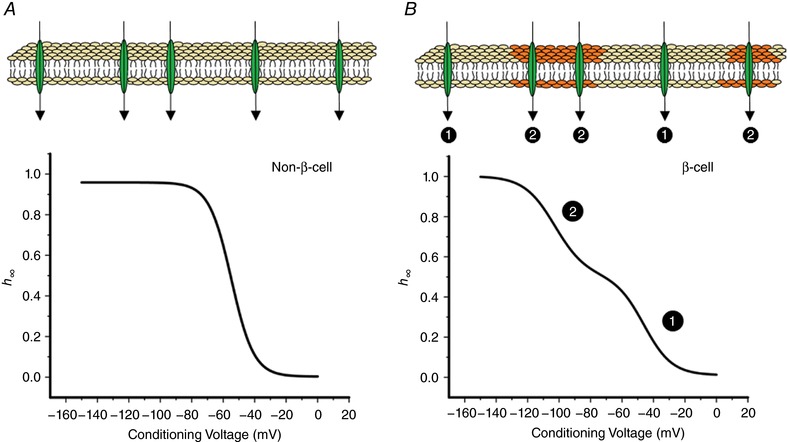
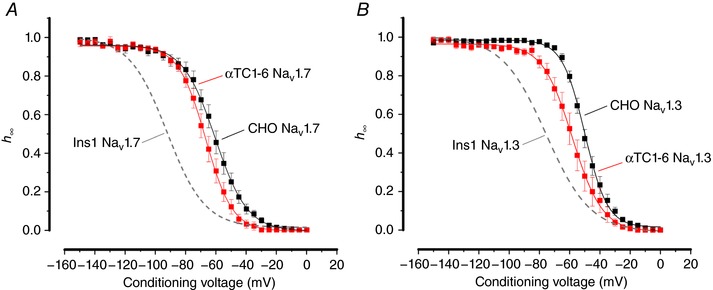
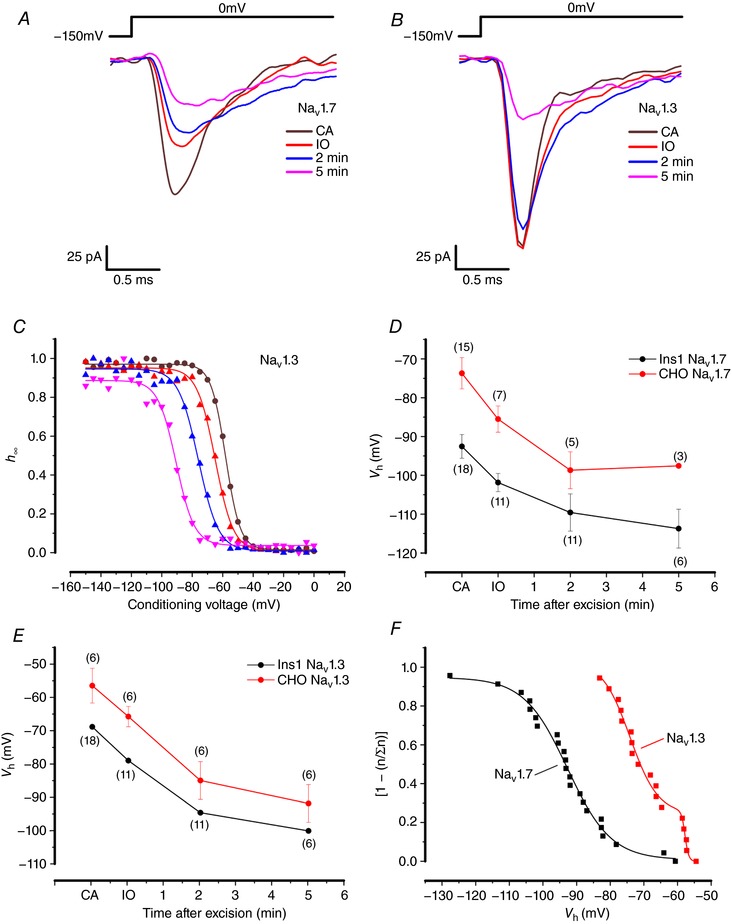
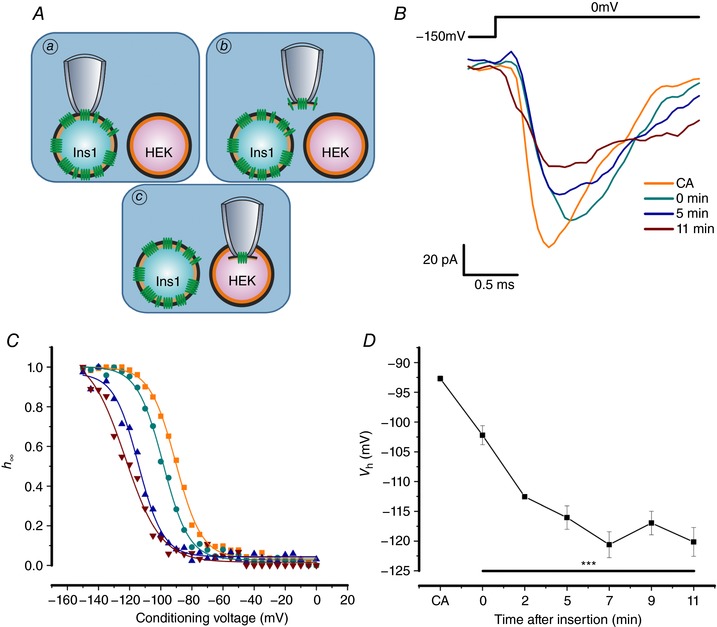
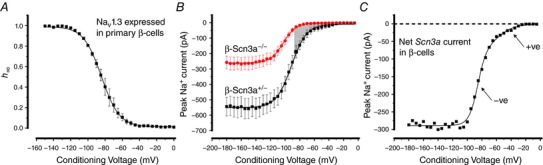
Na current inactivation is biphasic in insulin-secreting cells, proceeding with two voltage dependences that are half-maximal at ∼-100 mV and -60 mV. Inactivation of voltage-gated Na (Na ) channels occurs at ∼30 mV more negative voltages in insulin-secreting Ins1 and primary β-cells than in HEK, CHO or glucagon-secreting αTC1-6 cells. The difference in inactivation between Ins1 and non-β-cells persists in the inside-out patch configuration, discounting an involvement of a diffusible factor. In Ins1 cells and primary β-cells, but not in HEK cells, inactivation of a single Na subtype is biphasic and follows two voltage dependences separated by 30-40 mV. We propose that Na channels adopt different inactivation behaviours depending on the local membrane environment.
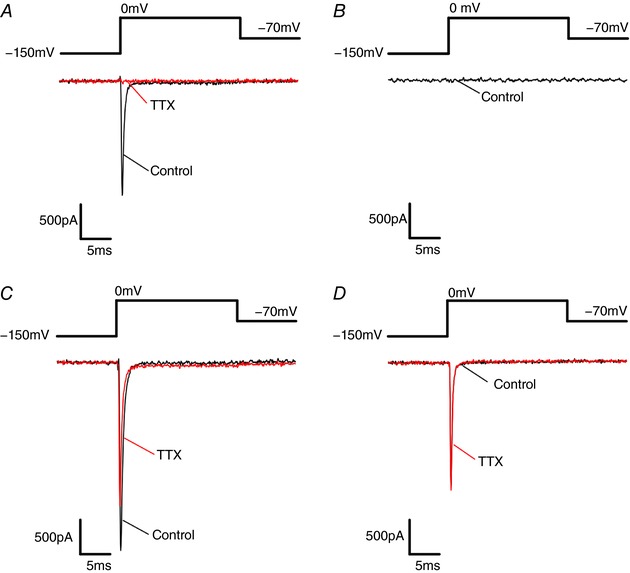
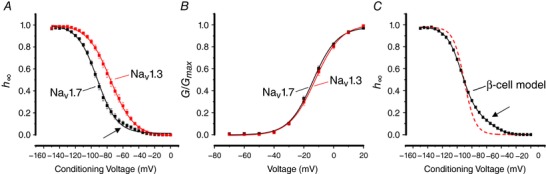
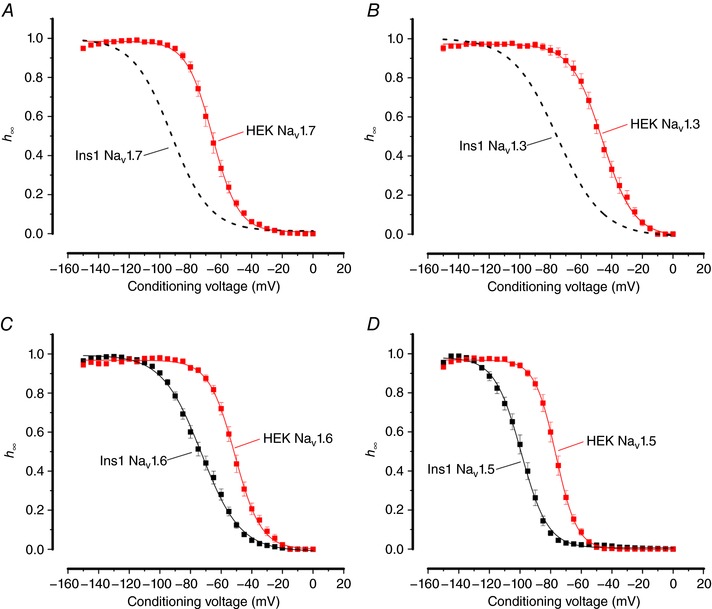
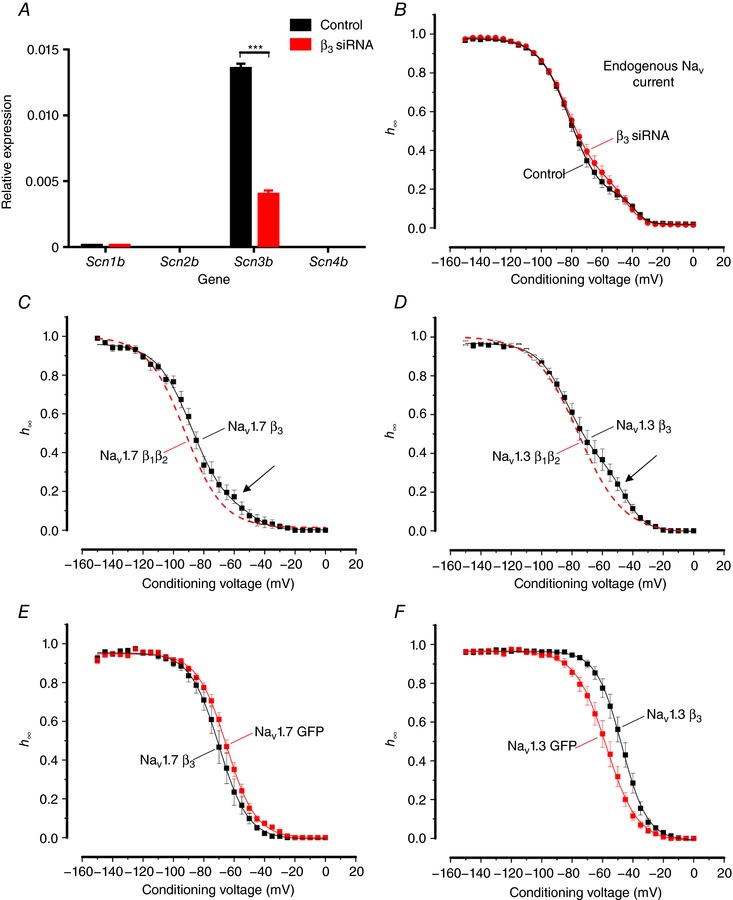
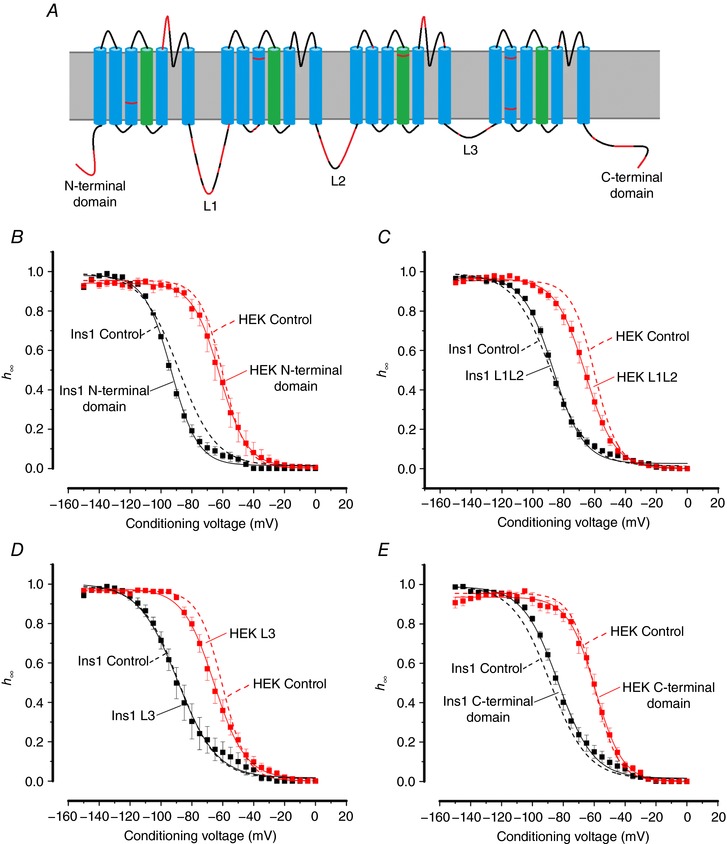
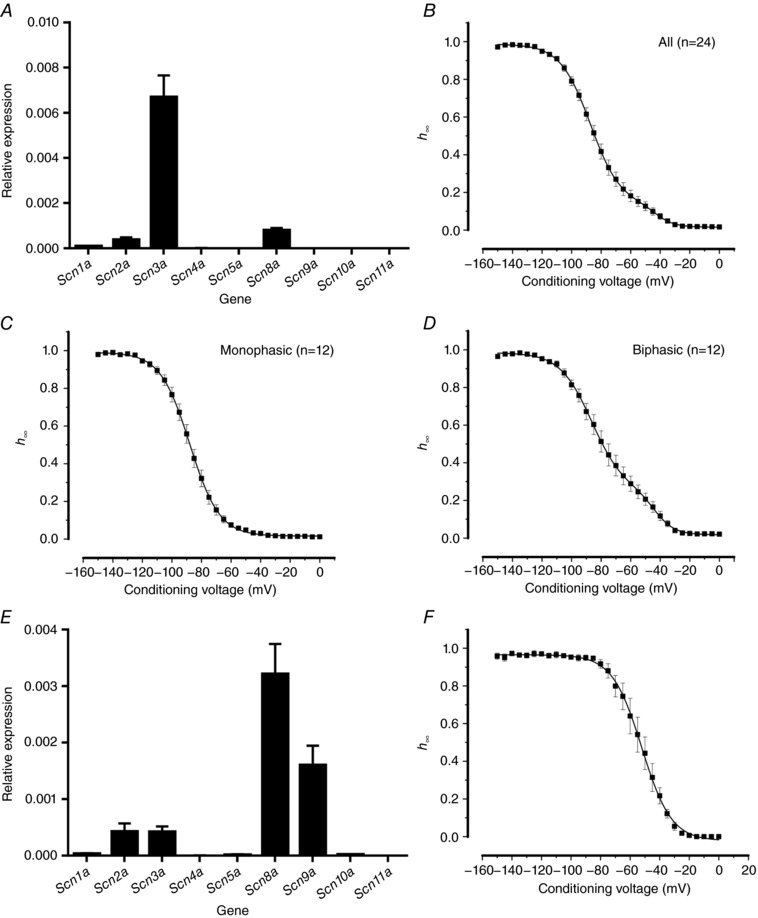
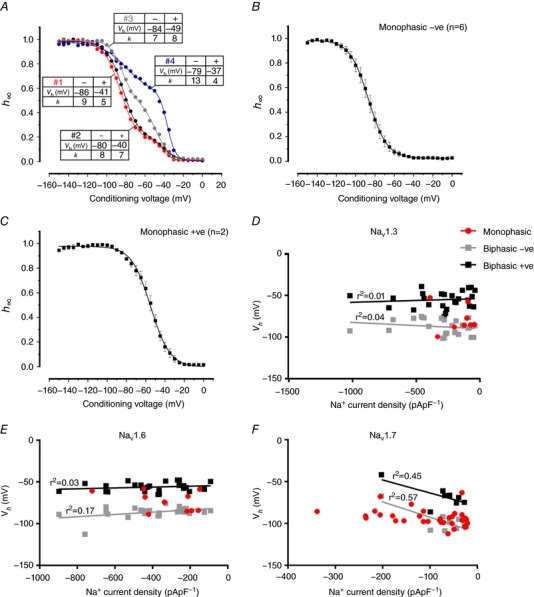
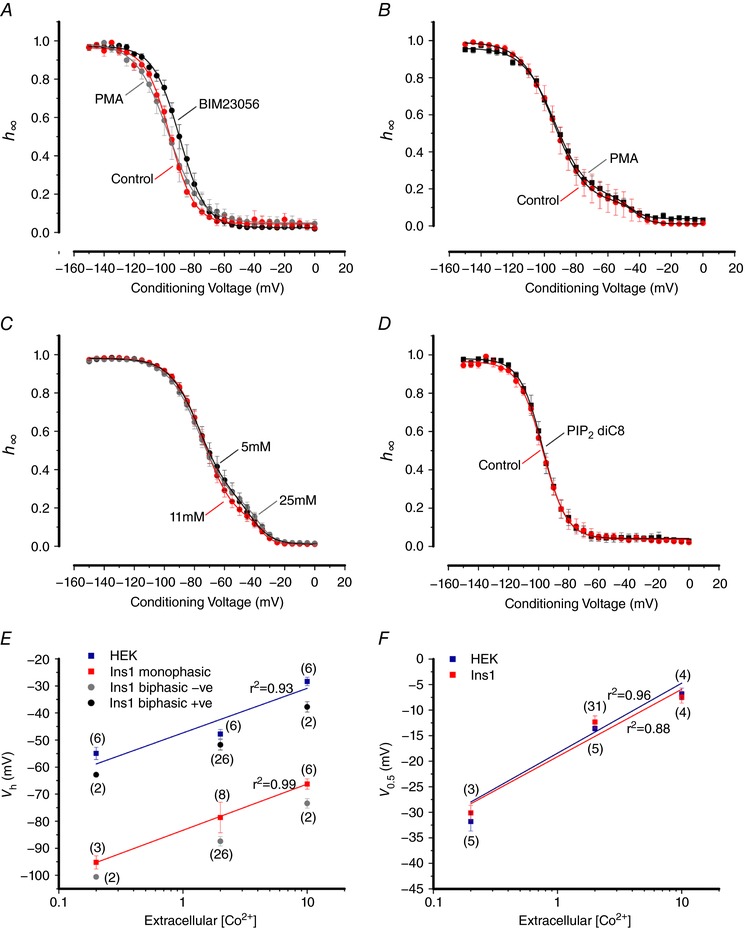
Pancreatic β-cells are equipped with voltage-gated Na channels that undergo biphasic voltage-dependent steady-state inactivation. A small Na current component (10-15%) inactivates over physiological membrane potentials and contributes to action potential firing. However, the major Na channel component is completely inactivated at -90 to -80 mV and is therefore inactive in the β-cell. It has been proposed that the biphasic inactivation reflects the contribution of different Na α-subunits. We tested this possibility by expression of TTX-resistant variants of the Na subunits found in β-cells (Na 1.3, Na 1.6 and Na 1.7) in insulin-secreting Ins1 cells and in non-β-cells (including HEK and CHO cells). We found that all Na subunits inactivated at 20-30 mV more negative membrane potentials in Ins1 cells than in HEK or CHO cells. The more negative inactivation in Ins1 cells does not involve a diffusible intracellular factor because the difference between Ins1 and CHO persisted after excision of the membrane. Na 1.7 inactivated at 15--20 mV more negative membrane potentials than Na 1.3 and Na 1.6 in Ins1 cells but this small difference is insufficient to solely explain the biphasic inactivation in Ins1 cells. In Ins1 cells, but never in the other cell types, widely different components of Na inactivation (separated by 30 mV) were also observed following expression of a single type of Na α-subunit. The more positive component exhibited a voltage dependence of inactivation similar to that found in HEK and CHO cells. We propose that biphasic Na inactivation in insulin-secreting cells reflects insertion of channels in membrane domains that differ with regard to lipid and/or membrane protein composition.
胰岛素分泌细胞中的钠电流失活呈双相性,具有两个电压依赖性,在约-100 mV 和-60 mV 时达到半最大值。与胰岛素分泌的 Ins1 和原代β细胞相比,电压门控钠(Na)通道在-30 mV 更负的电压下失活于 HEK、CHO 或胰高血糖素分泌的αTC1-6 细胞。Ins1 细胞和原代β细胞中失活的差异在内外位膜片钳配置中持续存在,排除了可扩散因子的参与。在 Ins1 细胞和原代β细胞中,但不在 HEK 细胞中,单一 Na 亚型的失活呈双相性,并遵循两个电压依赖性,由 30-40 mV 隔开。我们提出,Na 通道根据局部膜环境采用不同的失活行为。
胰腺β细胞配备有电压门控 Na 通道,该通道经历双相电压依赖性稳态失活。在生理膜电位下,小的 Na 电流成分(10-15%)失活并有助于动作电位发射。然而,主要的 Na 通道成分在-90 至-80 mV 完全失活,因此在β细胞中不活跃。有人提出,双相失活反映了不同 Naα 亚基的贡献。我们通过在胰岛素分泌的 Ins1 细胞和非β细胞(包括 HEK 和 CHO 细胞)中表达在β细胞中发现的 Na 亚基(Na1.3、Na1.6 和 Na1.7)的 TTX 抗性变体来检验这种可能性。我们发现,所有 Na 亚基在 Ins1 细胞中的失活比在 HEK 或 CHO 细胞中的负膜电位更负。Ins1 细胞中更负的失活不涉及可扩散的细胞内因子,因为在膜片钳切除后,Ins1 和 CHO 之间的差异仍然存在。在 Ins1 细胞中,Na1.7 的失活比 Na1.3 和 Na1.6 更负的膜电位,但这种小差异不足以单独解释 Ins1 细胞中的双相失活。在 Ins1 细胞中,但从未在其他细胞类型中,也观察到表达单个 Naα 亚基后,Na 失活的差异很大(相差 30 mV)。更正的成分表现出与在 HEK 和 CHO 细胞中发现的失活电压依赖性相似。我们提出,胰岛素分泌细胞中双相 Na 失活反映了通道插入脂质和/或膜蛋白组成不同的膜域。