Federal State Budget Scientific Institution "The Federal Research Center Institute of Cytology and Genetics of Siberian Branch of the Russian Academy of Sciences", Novosibirsk, Russia.
Novosibirsk State University, Novosibirsk, Russia.
BMC Genomics. 2018 Feb 9;19(Suppl 3):114. doi: 10.1186/s12864-018-4473-8.
MicroRNAs proceeds through the different canonical and non-canonical pathways; the most frequent of the non-canonical ones is the splicing-dependent biogenesis of mirtrons. We compare the mirtrons and non-mirtrons of human and mouse to explore how their maturation appears in the precursor structure around the miRNA.
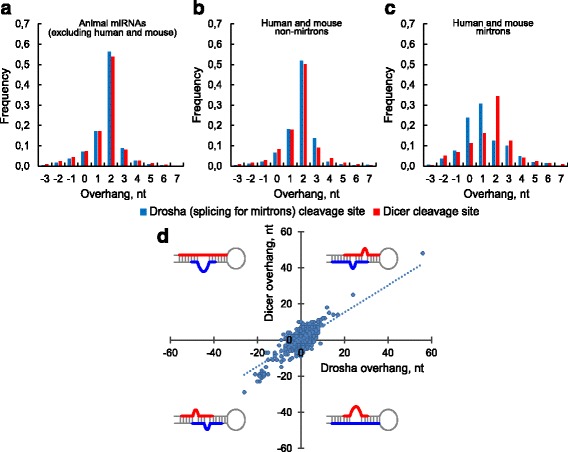
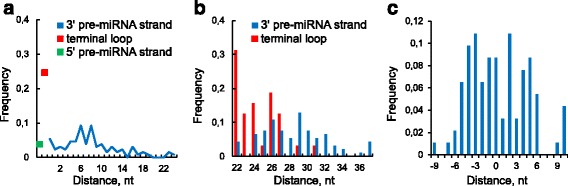
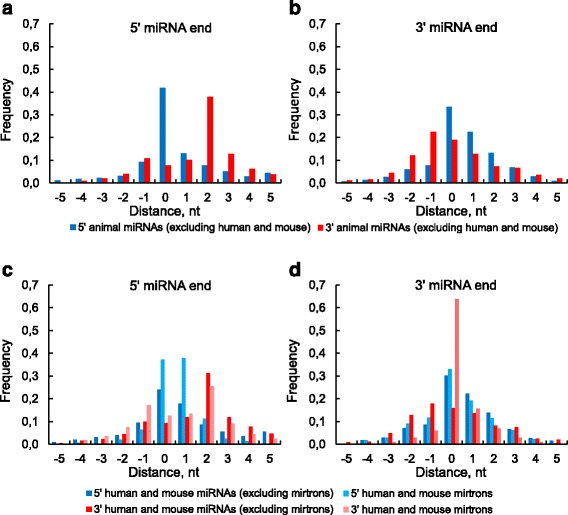
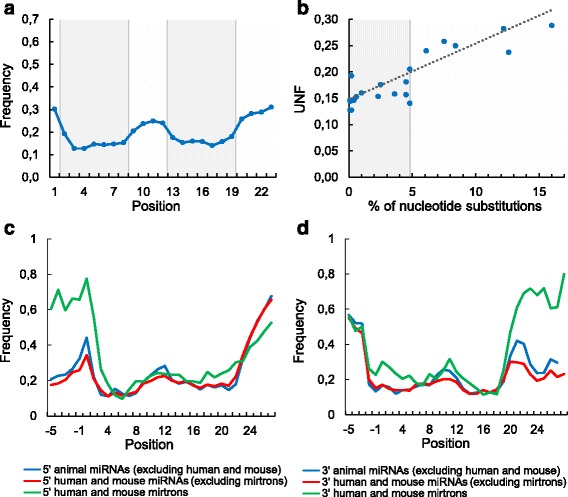
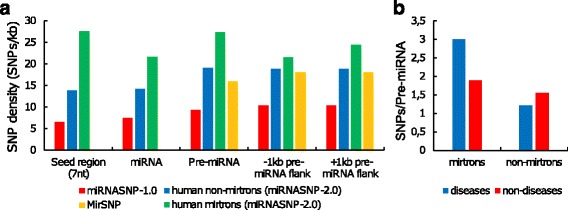
We found the coherence of the overhang lengths what indicates the dependence between the cleavage sites. To explain this dependence we suggest the 2-lever model of the Dicer structure that couples the imprecisions in Drosha and Dicer. Considering the secondary structure of all animal pre-miRNAs we confirmed that single-stranded nucleotides tend to be located near the miRNA boundaries and in its center and are characterized by a higher mutation rate. The 5' end of the canonical 5' miRNA approaches the nearest single-stranded nucleotides what suggests the extension of the loop-counting rule from the Dicer to the Drosha cleavage site. A typical structure of the annotated mirtron pre-miRNAs differs from the canonical pre-miRNA structure and possesses the 1- and 2 nt hanging ends at the hairpin base. Together with the excessive variability of the mirtron Dicer cleavage site (that could be partially explained by guanine at its ends inherited from splicing) this is one more evidence for the 2-lever model. In contrast with the canonical miRNAs the mirtrons have higher snp densities and their pre-miRNAs are inversely associated with diseases. Therefore we supported the view that mirtrons are under positive selection while canonical miRNAs are under negative one and we suggested that mirtrons are an intrinsic source of silencing variability which produces the disease-promoting variants. Finally, we considered the interference of the pre-miRNA structure and the U2snRNA:pre-mRNA basepairing. We analyzed the location of the branchpoints and found that mirtron structure tends to expose the branchpoint site what suggests that the mirtrons can readily evolve from occasional hairpins in the immediate neighbourhood of the 3' splice site.
The miRNA biogenesis manifests itself in the footprints of the secondary structure. Close inspection of these structural properties can help to uncover new pathways of miRNA biogenesis and to refine the known miRNA data, in particular, new non-canonical miRNAs may be predicted or the known miRNAs can be re-classified.
microRNAs 通过不同的经典和非经典途径进行加工;最常见的非经典途径是剪接依赖的 mirtron 生物发生。我们比较了人和小鼠的 mirtron 和非 mirtron,以探索它们的成熟如何出现在 miRNA 周围的前体结构中。
我们发现了侧翼长度的一致性,这表明切割位点之间存在依赖性。为了解释这种依赖性,我们提出了 Dicer 结构的双杠杆模型,该模型将 Drosha 和 Dicer 的不精确性联系起来。考虑到所有动物前 miRNA 的二级结构,我们证实单链核苷酸倾向于位于 miRNA 边界附近及其中心,并且具有更高的突变率。经典 5' miRNA 的 5' 端接近最近的单链核苷酸,这表明从 Dicer 到 Drosha 切割位点的环计数规则的扩展。注释的 mirtron 前 miRNA 的典型结构与经典前 miRNA 结构不同,在发夹基底部具有 1 个和 2 个核苷酸的悬垂末端。与 mirtron Dicer 切割位点的过度可变性(其末端的鸟嘌呤可能部分由剪接继承)一起,这是双杠杆模型的另一个证据。与经典 miRNA 相比,mirtrons 具有更高的 SNP 密度,并且它们的前 miRNA 与疾病呈负相关。因此,我们支持这样的观点,即 mirtrons 受到正选择,而经典 miRNA 受到负选择,我们还提出 mirtrons 是沉默变异性的内在来源,产生促进疾病的变体。最后,我们考虑了前 miRNA 结构和 U2snRNA:pre-mRNA 碱基配对的干扰。我们分析了分支点的位置,发现 mirtron 结构倾向于暴露分支点位点,这表明 mirtrons 可以很容易地从 3' 剪接位点附近偶尔的发夹中进化而来。
miRNA 的生物发生表现在二级结构的足迹中。仔细检查这些结构特性可以帮助揭示 miRNA 生物发生的新途径,并完善已知的 miRNA 数据,特别是可以预测新的非经典 miRNA 或对已知 miRNA 进行重新分类。