National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Alaska Fisheries Science Center, Seattle, WA 98115;
Quantitative Ecology and Resource Management, University of Washington, Seattle, WA 98195.
Proc Natl Acad Sci U S A. 2018 May 8;115(19):4945-4950. doi: 10.1073/pnas.1800096115. Epub 2018 Apr 19.
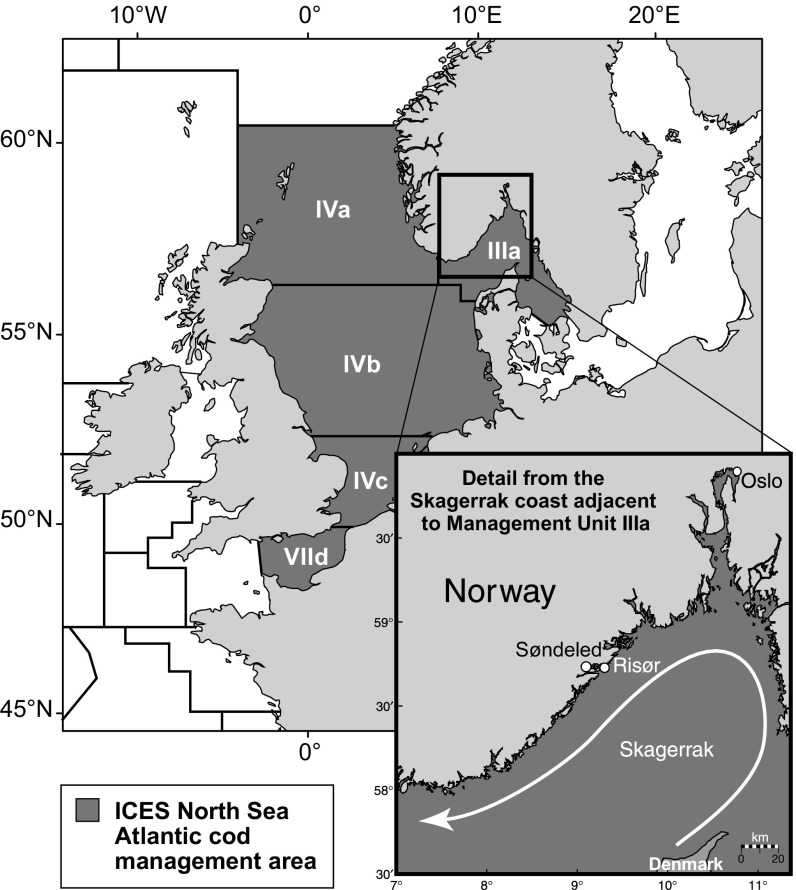
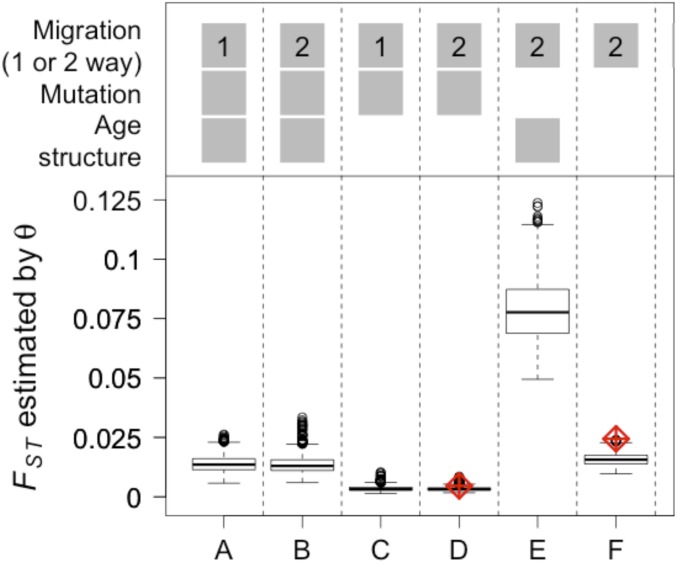
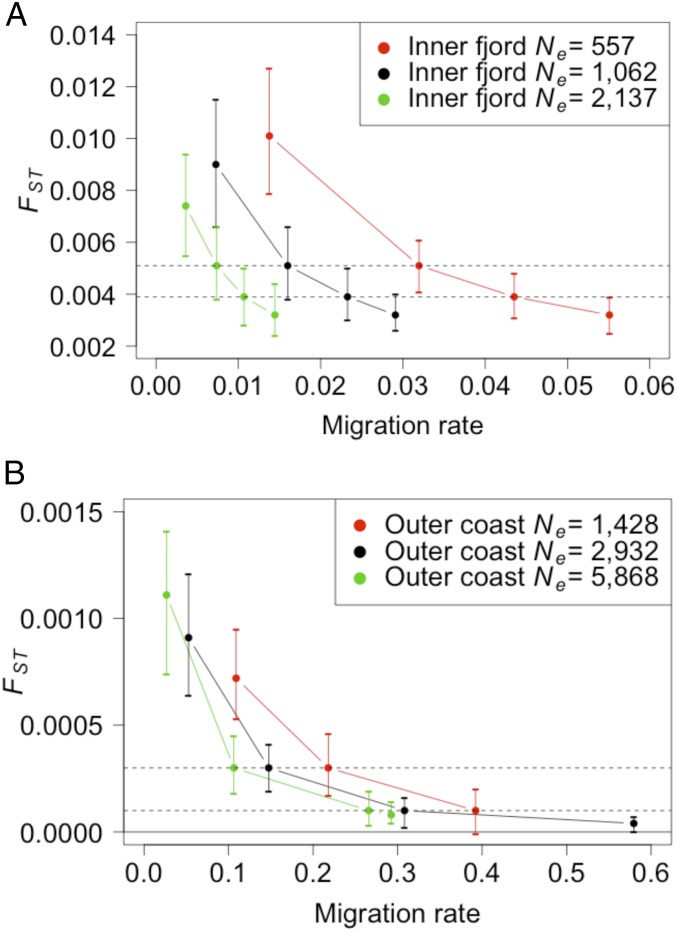
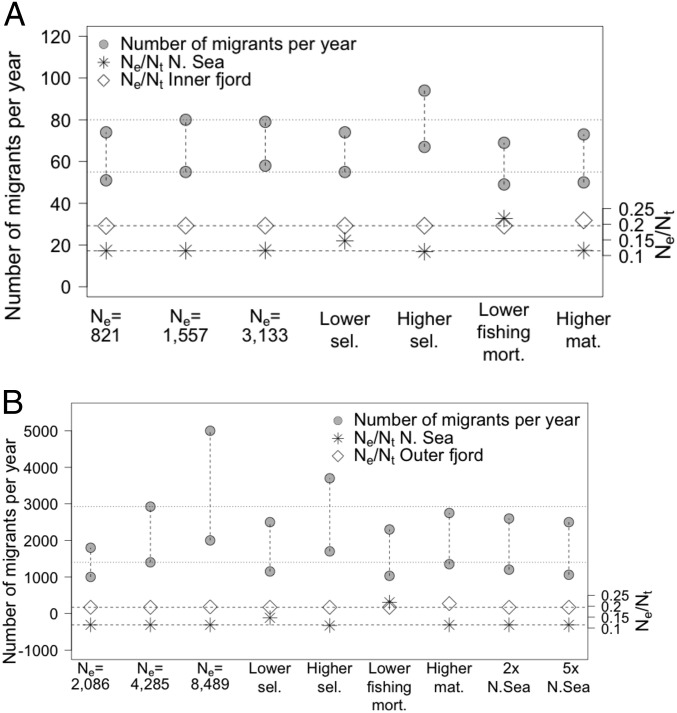
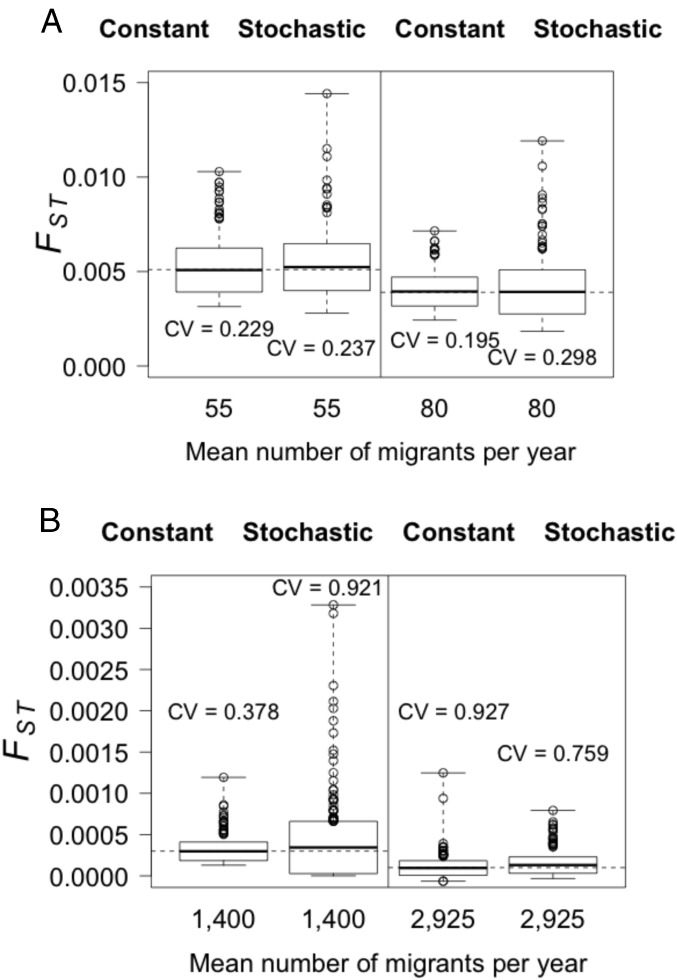
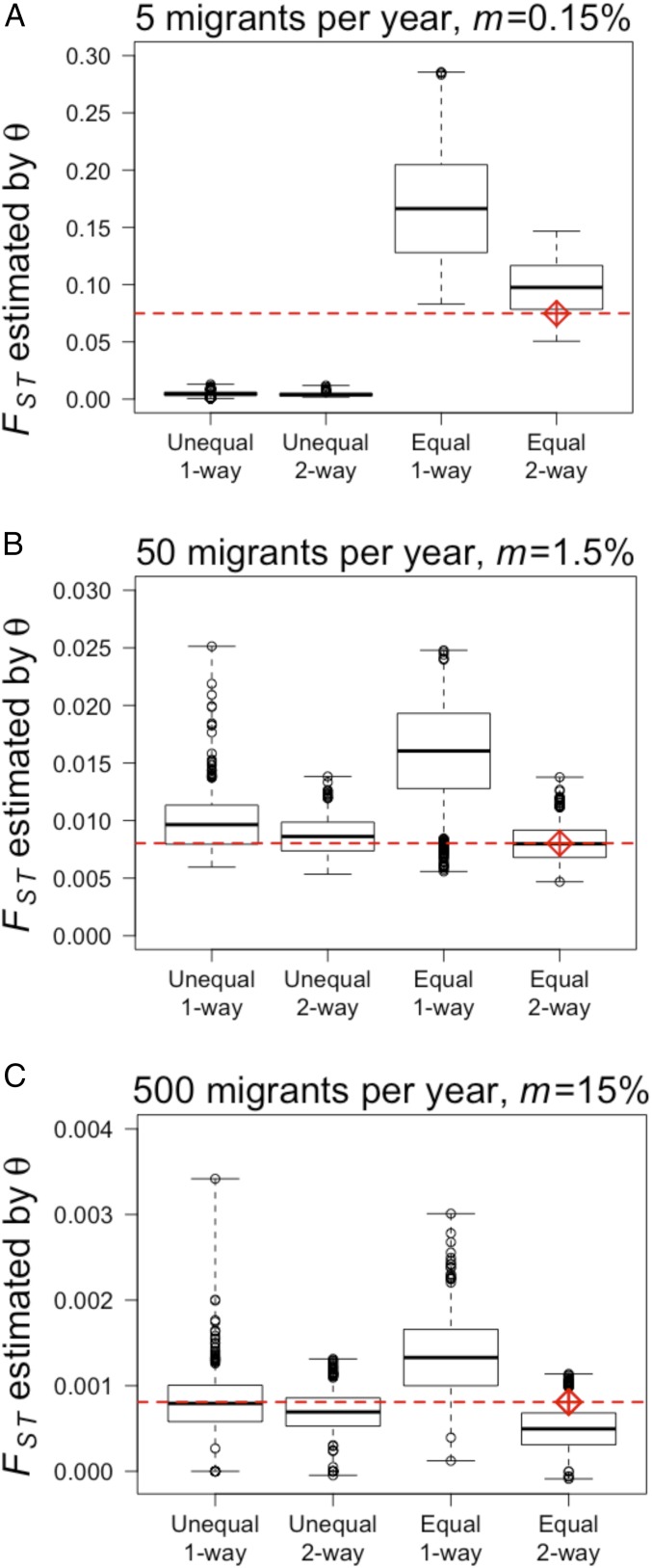
Genetic data are commonly used to estimate connectivity between putative populations, but translating them to demographic dispersal rates is complicated. Theoretical equations that infer a migration rate based on the genetic estimator , such as Wright's equation, ≈ 1/(4 + 1), make assumptions that do not apply to most real populations. How complexities inherent to real populations affect migration was exemplified by Atlantic cod in the North Sea and Skagerrak and was examined within an age-structured model that incorporated genetic markers. Migration was determined under various scenarios by varying the number of simulated migrants until the mean simulated level of genetic differentiation matched a fixed level of genetic differentiation equal to empirical estimates. Parameters that decreased the / ratio (where is the effective and is the total population size), such as high fishing mortality and high fishing gear selectivity, increased the number of migrants required to achieve empirical levels of genetic differentiation. Higher maturity-at-age and lower selectivity increased / and decreased migration when genetic differentiation was fixed. Changes in natural mortality, fishing gear selectivity, and maturity-at-age within expected limits had a moderate effect on migration when genetic differentiation was held constant. Changes in population size had the greatest effect on the number of migrants to achieve fixed levels of , particularly when genetic differentiation was low, ≈ 10 Highly variable migration patterns, compared with constant migration, resulted in higher variance in genetic differentiation and higher extreme values. Results are compared with and provide insight into the use of theoretical equations to estimate migration among real populations.
遗传数据通常用于估计假定群体之间的连通性,但将其转化为人口扩散率是复杂的。基于遗传估计器 (如 Wright 方程, ≈ 1/(4 + 1))推断迁移率的理论方程存在一些假设,这些假设并不适用于大多数真实种群。真实种群中固有的复杂性如何影响迁移,以北海和斯卡格拉克的大西洋鳕鱼为例,并在一个包含遗传标记的年龄结构模型中进行了检验。通过改变模拟移民的数量,在各种情况下确定迁移,直到模拟的遗传分化平均值与固定的遗传分化水平相匹配,该水平等于经验估计值。降低 / 比(其中 是有效和 是总种群大小)的参数,如高捕捞死亡率和高渔具选择性,增加了实现经验遗传分化水平所需的移民数量。较高的年龄成熟度和较低的选择性增加了 / 并降低了遗传分化固定时的迁移。当遗传分化保持不变时,自然死亡率、渔具选择性和年龄成熟度的变化对迁移的影响适中。在种群规模发生变化时,对实现固定 水平的移民数量的影响最大,尤其是当遗传分化较低时, ≈ 10。与恒定迁移相比,高度可变的迁移模式导致遗传分化的方差更高和极值更高。结果与理论方程用于估计真实种群之间的迁移进行了比较,并提供了深入的了解。