Holub Daniel, Ma Hongju, Krauß Norbert, Lamparter Tilman, Elstner Marcus, Gillet Natacha
Department for Theoretical Chemical Biology , Institute for Physical Chemistry , Karlsruhe Institute for Technology , Kaiserstr. 12 , 76131 , Karlsruhe , Germany . Email:
Botanical Institute , Karlsruhe Institute for Technology , Fritz Haber Weg 4 , 76131 , Karlsruhe , Germany.
Chem Sci. 2017 Dec 11;9(5):1259-1272. doi: 10.1039/c7sc03386a. eCollection 2018 Feb 7.
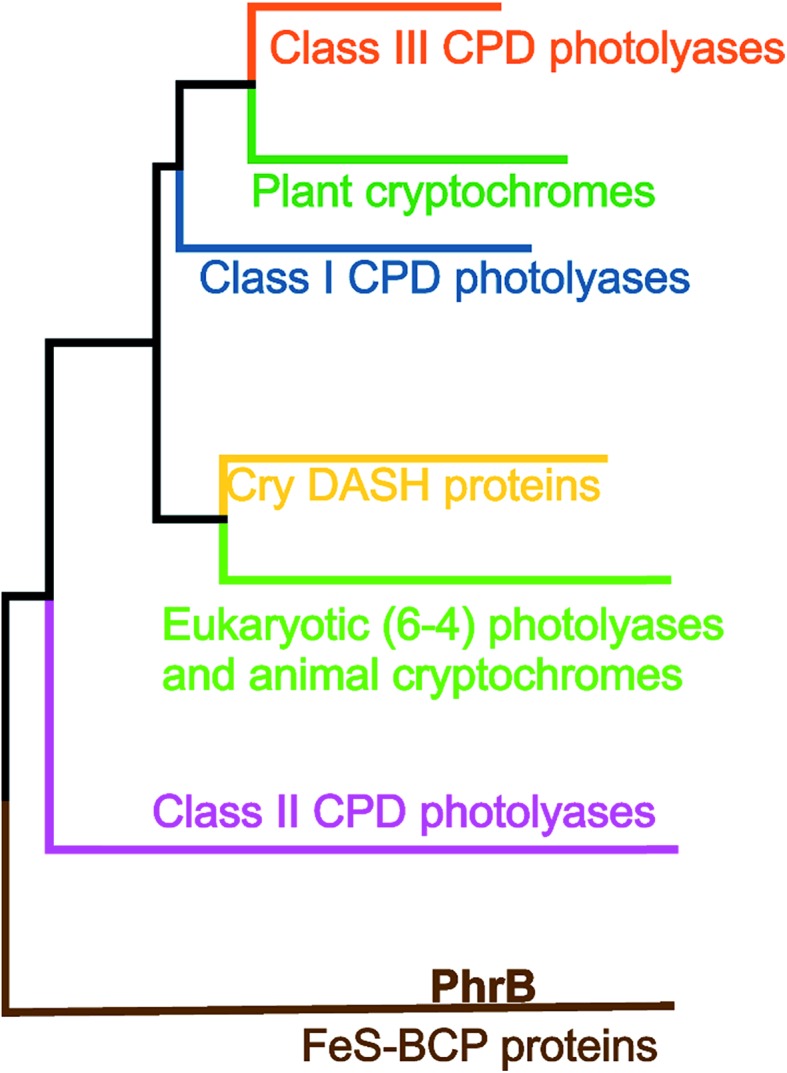
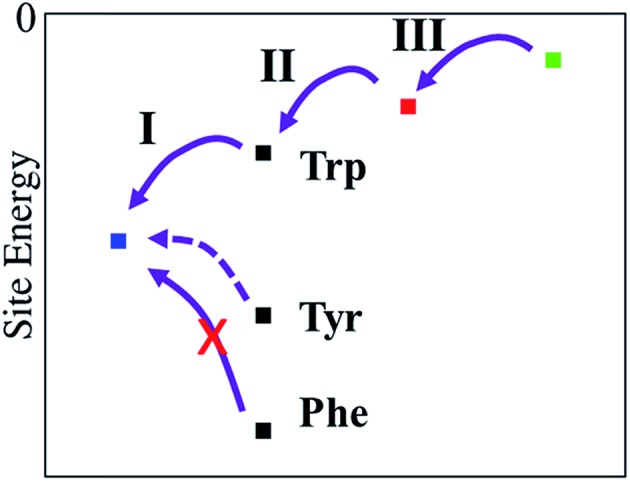
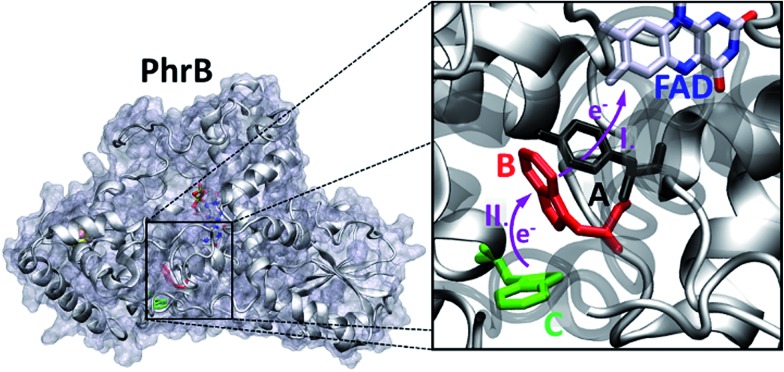
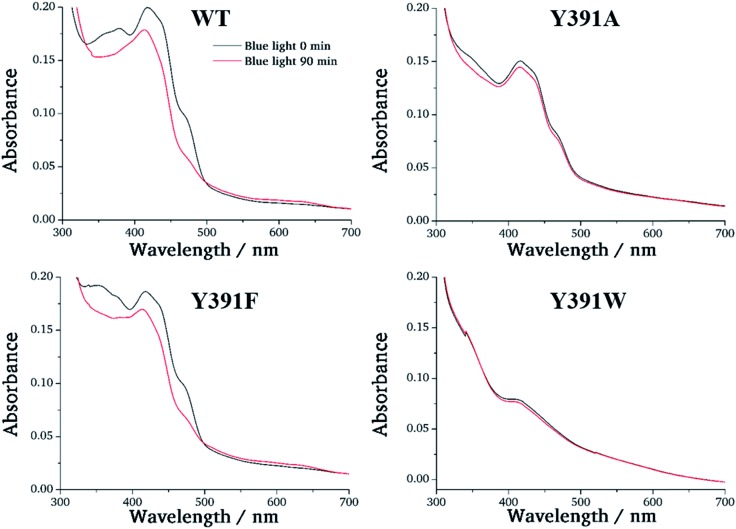
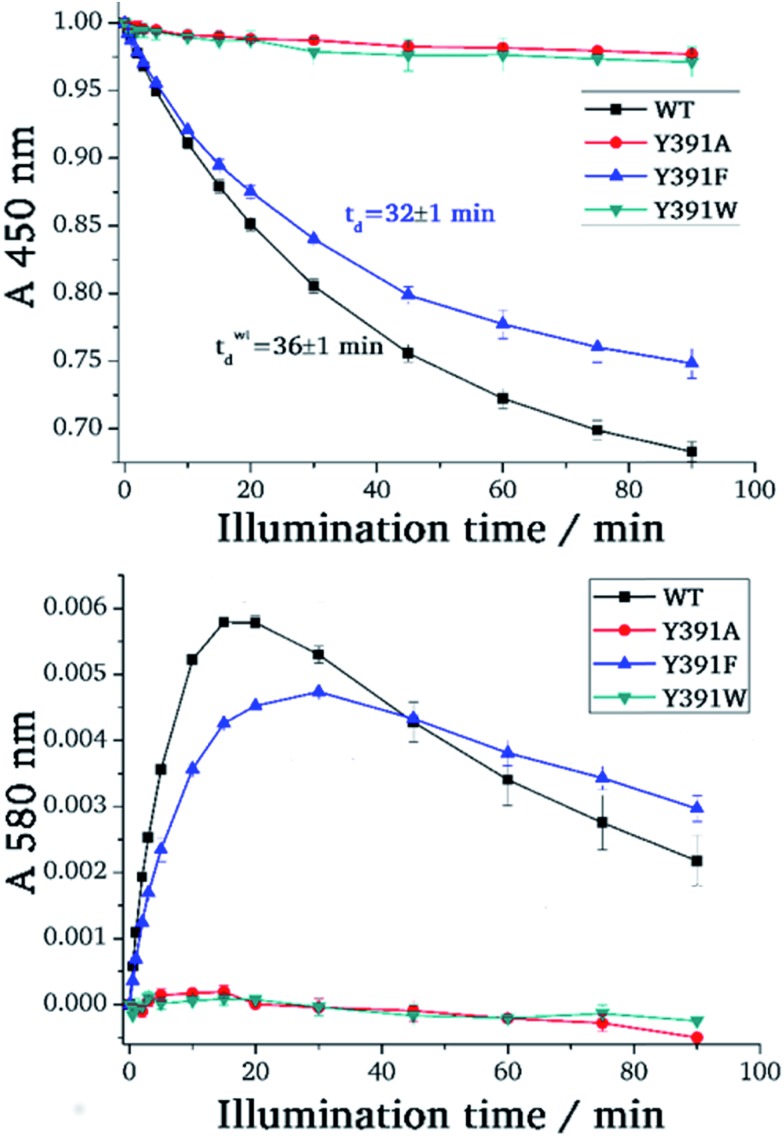
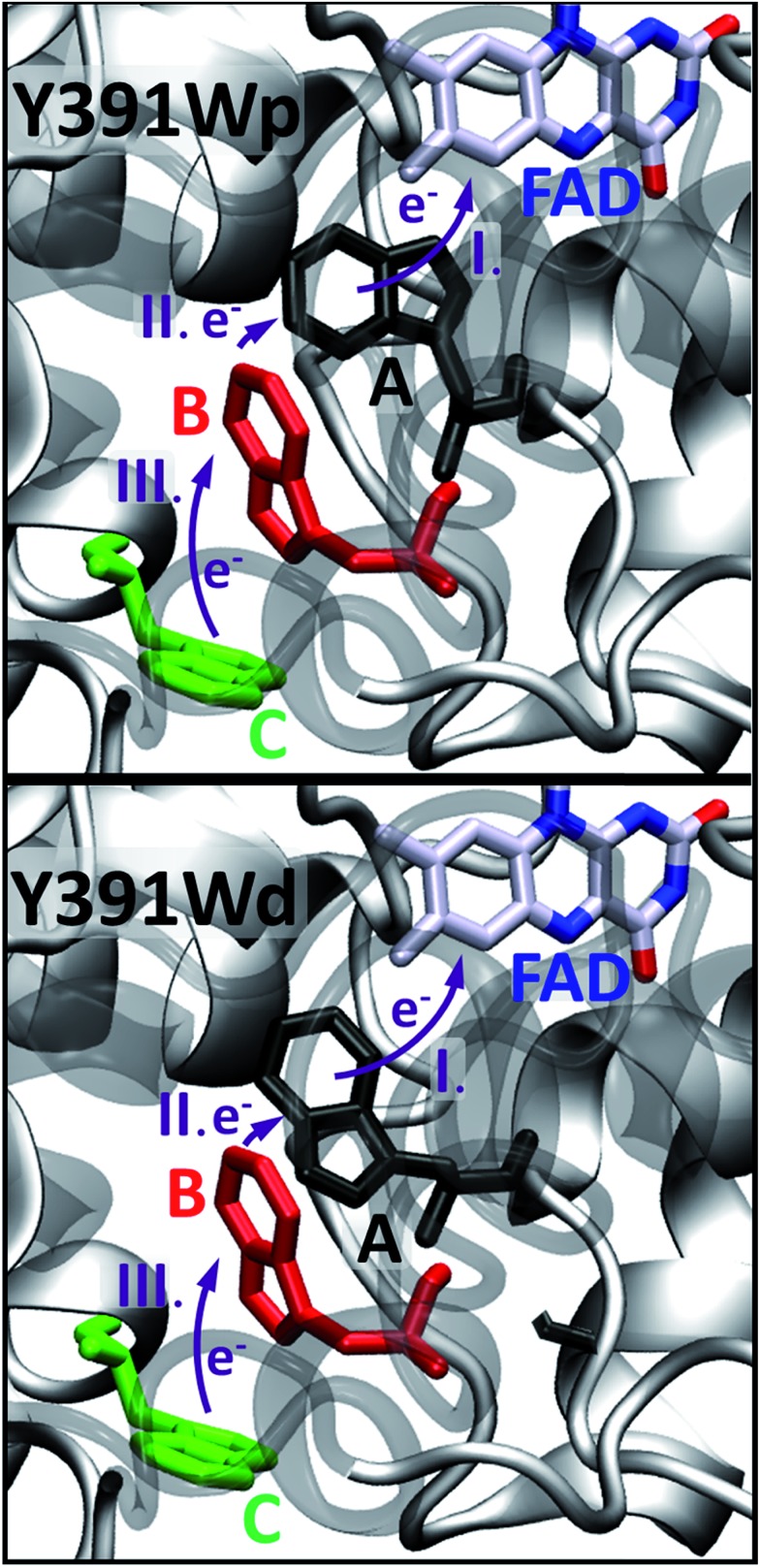
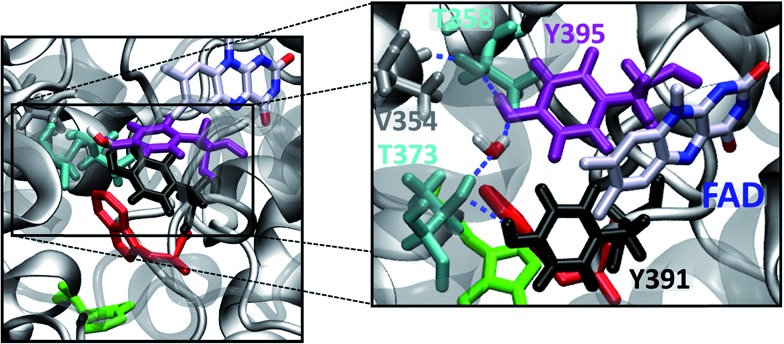
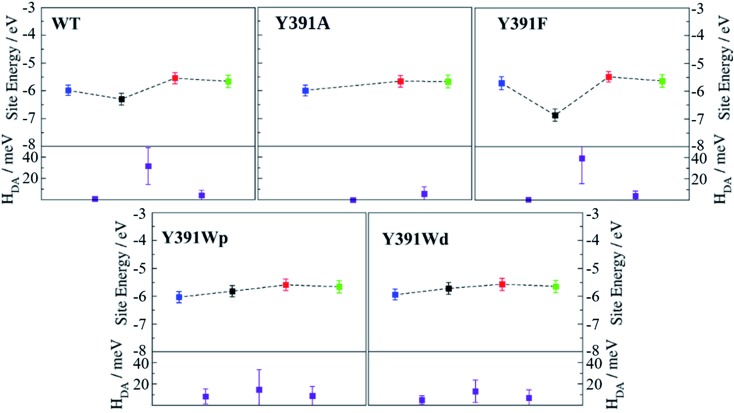
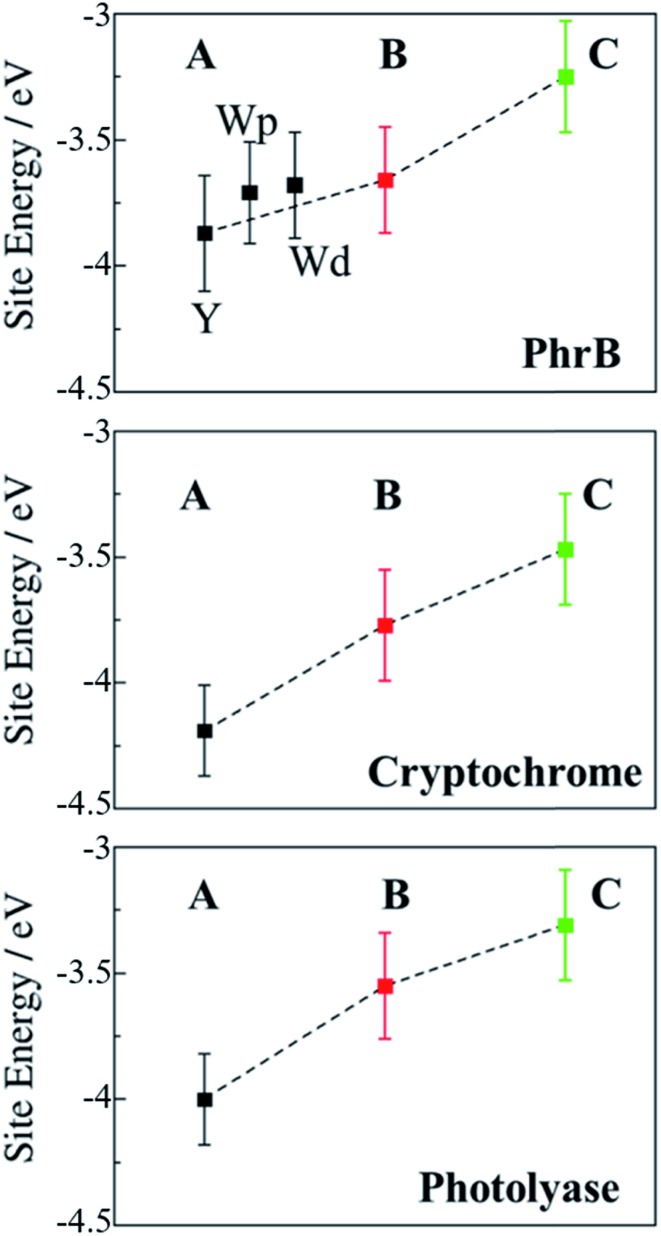
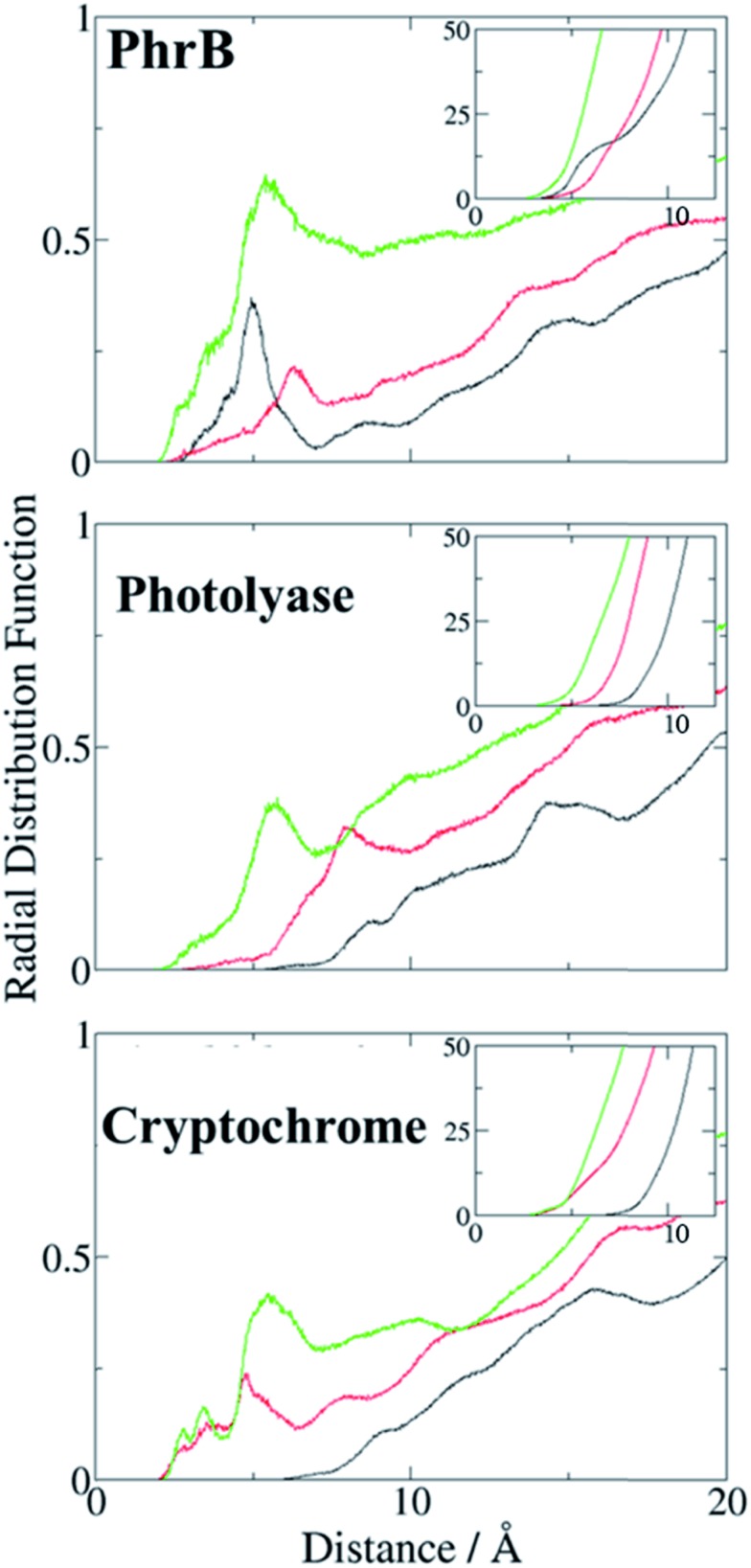
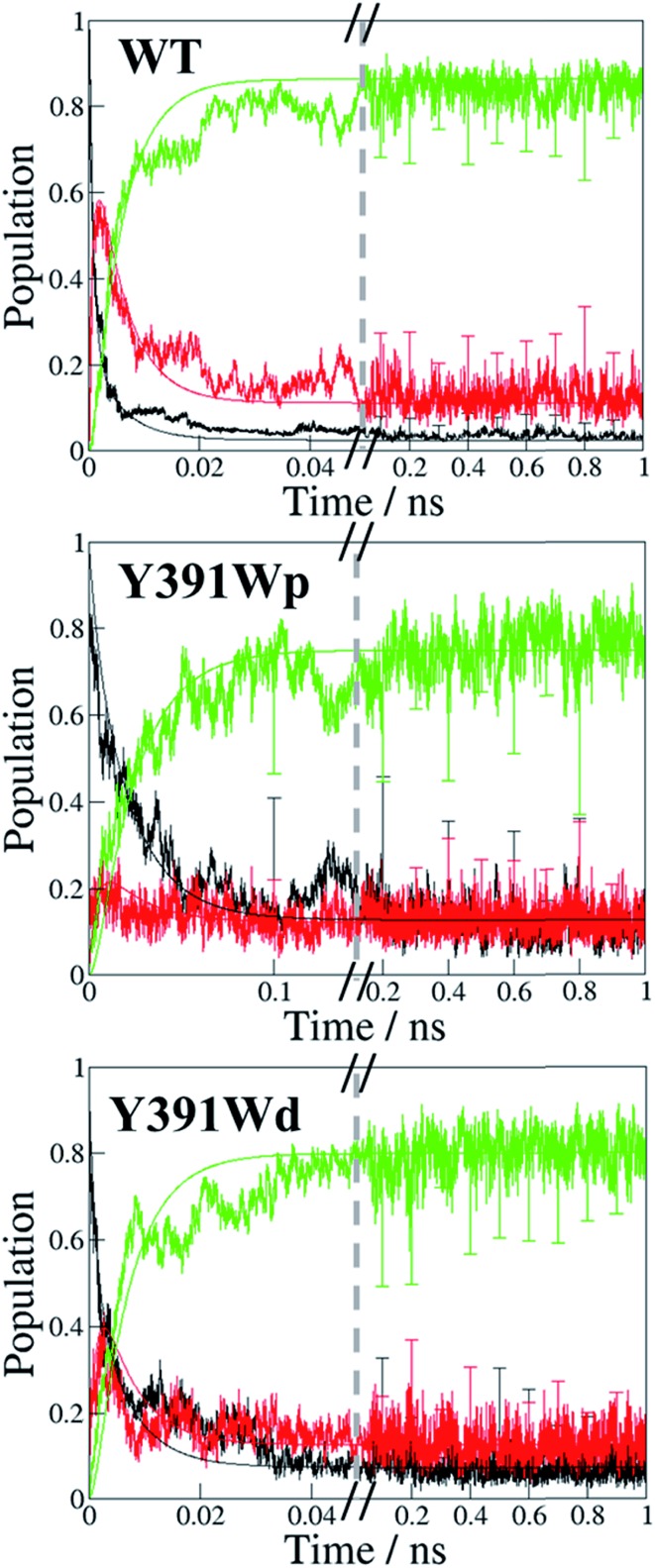
Cryptochromes and photolyases form a flavoprotein family in which the FAD chromophore undergoes light induced changes of its redox state. During this process, termed photoreduction, electrons flow from the surface conserved amino acid residues to FAD. The bacterial (6-4) photolyase PhrB belongs to a phylogenetically ancient group. Photoreduction of PhrB differs from the typical pattern because the amino acid of the electron cascade next to FAD is a tyrosine (Tyr391), whereas photolyases and cryptochromes of other groups have a tryptophan as direct electron donor of FAD. Mutagenesis studies have identified Trp342 and Trp390 as essential for charge transfer. Trp342 is located at the periphery of PhrB while Trp390 connects Trp342 and Tyr391. The role of Tyr391, which lies between Trp390 and FAD, is however unclear as its replacement by phenylalanine did not block photoreduction. Experiments reported here, which replace Tyr391 by Ala, show that photoreduction is blocked, underlining the relevance of Tyr/Phe at position 391 and indicating that charge transfer occurs the triad 391-390-342. This raises the question, why PhrB positions a tyrosine at this location, having a less favourable ionisation potential than tryptophan, which occurs at this position in many proteins of the photolyase/cryptochrome family. Tunnelling matrix calculations show that tyrosine or phenylalanine can be involved in a productive bridged electron transfer between FAD and Trp390, in line with experimental findings. Since replacement of Tyr391 by Trp resulted in loss of FAD and DMRL chromophores, electron transfer cannot be studied experimentally in this mutant, but calculations on a mutant model suggest that Trp might participate in the electron transfer cascade. Charge transfer simulations reveal an unusual stabilization of the positive charge on site 391 compared to other photolyases or cryptochromes. Water molecules near Tyr391 offer a polar environment which stabilizes the positive charge on this site, thereby lowering the energetic barrier intrinsic to tyrosine. This opens a second charge transfer channel in addition to tunnelling through the tyrosine barrier, based on hopping and therefore transient oxidation of Tyr391, which enables a fast charge transfer similar to proteins utilizing a tryptophan-triad. Our results suggest that evolution of the first site of the redox chain has just been possible by tuning the protein structure and environment to manage a downhill hole transfer process from FAD to solvent.
隐花色素和光解酶构成一个黄素蛋白家族,其中黄素腺嘌呤二核苷酸(FAD)发色团会经历光诱导的氧化还原状态变化。在这个被称为光还原的过程中,电子从表面保守的氨基酸残基流向FAD。细菌(6 - 4)光解酶PhrB属于一个系统发育上古老的群体。PhrB的光还原不同于典型模式,因为FAD旁边电子传递链中的氨基酸是酪氨酸(Tyr391),而其他群体的光解酶和隐花色素有一个色氨酸作为FAD的直接电子供体。诱变研究已确定Trp342和Trp390对电荷转移至关重要。Trp342位于PhrB的外围,而Trp390连接Trp342和Tyr391。然而,位于Trp390和FAD之间的Tyr391的作用尚不清楚,因为用苯丙氨酸取代它并没有阻止光还原。本文报道的用丙氨酸取代Tyr391的实验表明光还原被阻断,强调了391位酪氨酸/苯丙氨酸的相关性,并表明电荷转移发生在391 - 390 - 342三联体中。这就提出了一个问题,为什么PhrB在这个位置定位一个酪氨酸,其电离势比色氨酸更不利,而色氨酸在光解酶/隐花色素家族的许多蛋白质的这个位置都存在。隧道矩阵计算表明,酪氨酸或苯丙氨酸可以参与FAD和Trp390之间有效的桥连电子转移,这与实验结果一致。由于用色氨酸取代Tyr391导致FAD和DMRL发色团丢失,无法在这个突变体中通过实验研究电子转移,但对突变体模型的计算表明色氨酸可能参与电子传递链。电荷转移模拟显示,与其他光解酶或隐花色素相比,391位点的正电荷有异常的稳定。Tyr391附近的水分子提供了一个极性环境,稳定了该位点的正电荷,从而降低了酪氨酸固有的能量屏障。这除了通过酪氨酸屏障的隧道效应外,还开辟了第二个电荷转移通道,该通道基于跳跃以及因此Tyr391的瞬时氧化,使得电荷转移速度与利用色氨酸三联体的蛋白质相似。我们的结果表明,通过调整蛋白质结构和环境以管理从FAD到溶剂的下坡空穴转移过程,氧化还原链第一个位点的进化才成为可能。