Hasan Cris R, Krauskopf Bernd, Osinga Hinke M
Department of Mathematics, The University of Auckland, Auckland, New Zealand.
J Math Neurosci. 2018 Apr 19;8(1):5. doi: 10.1186/s13408-018-0060-1.
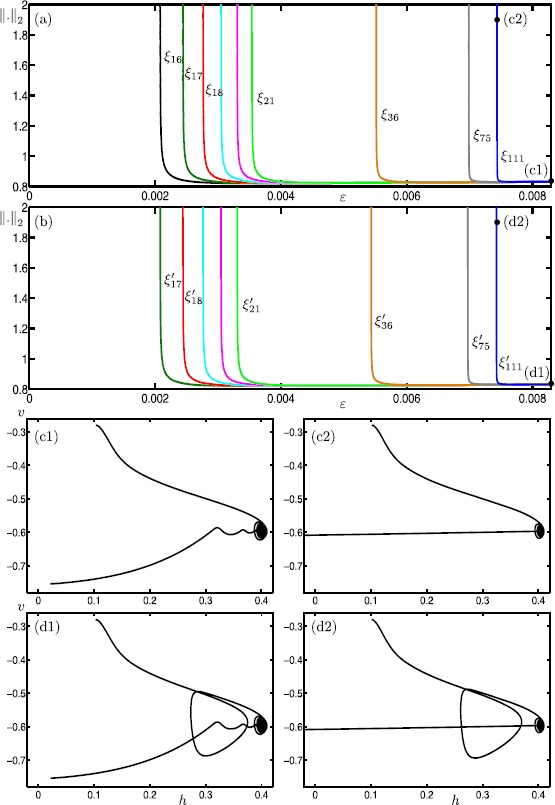
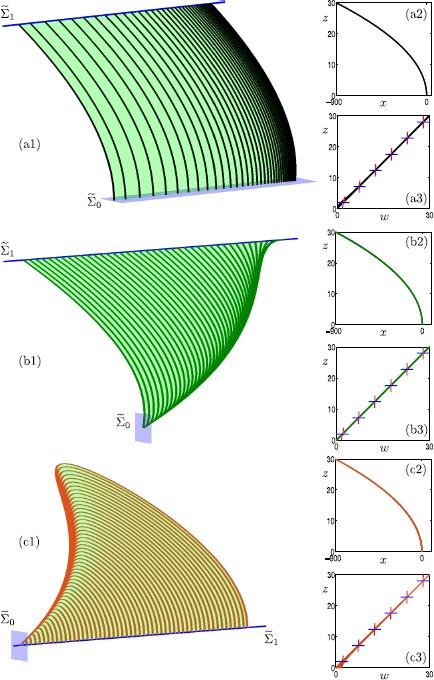
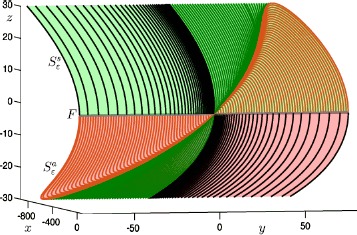
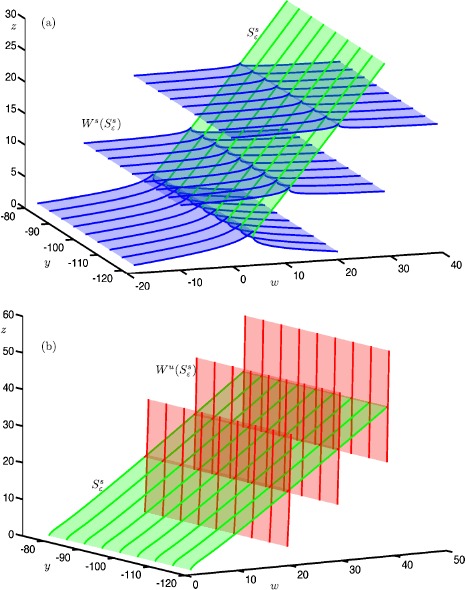
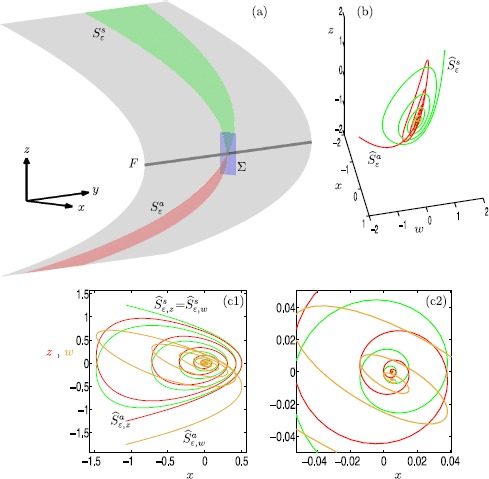
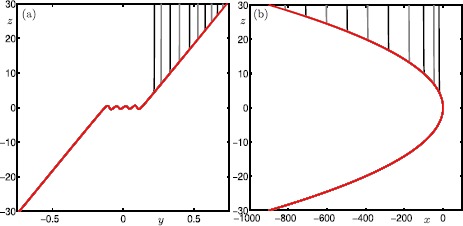
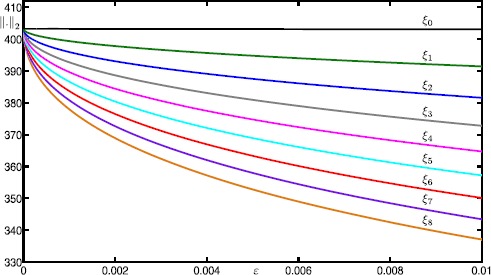
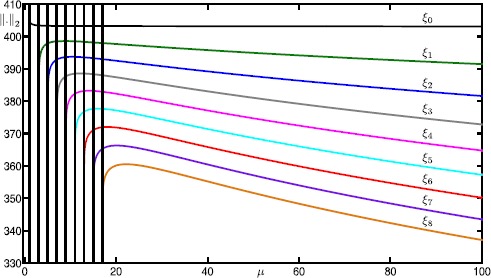
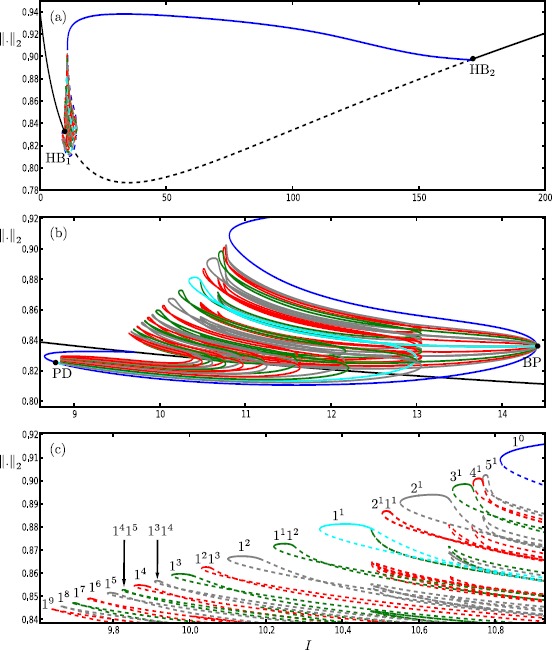
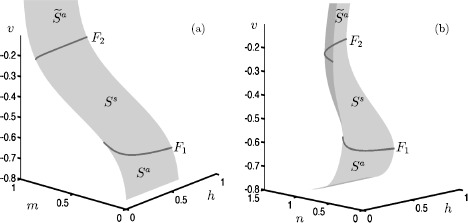
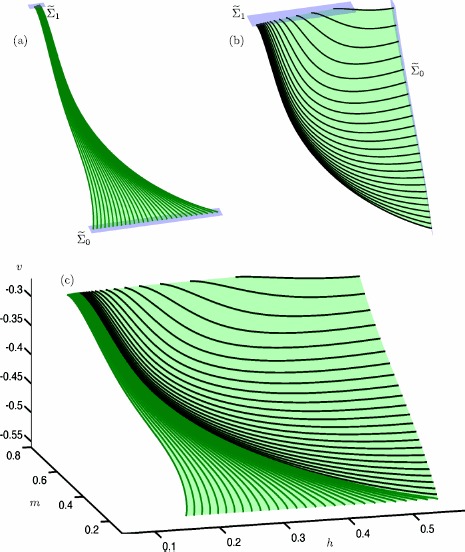
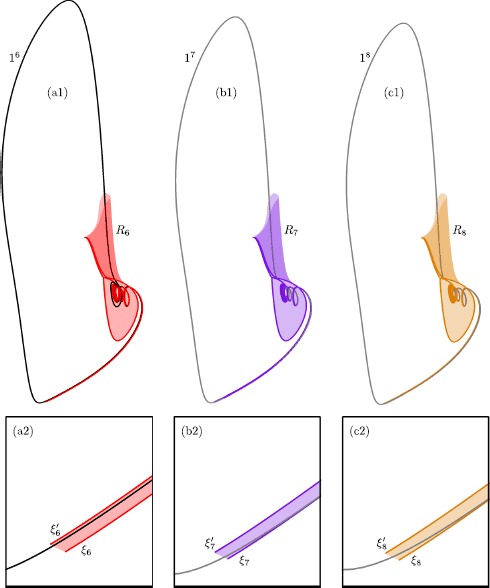
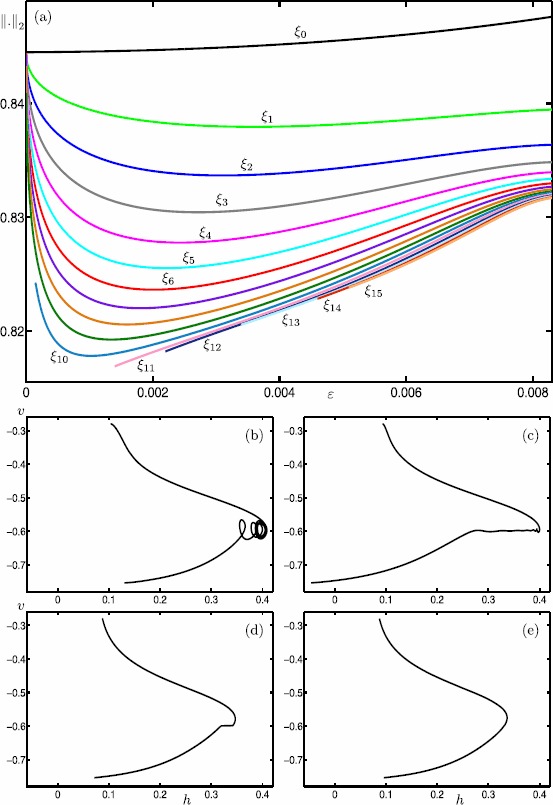
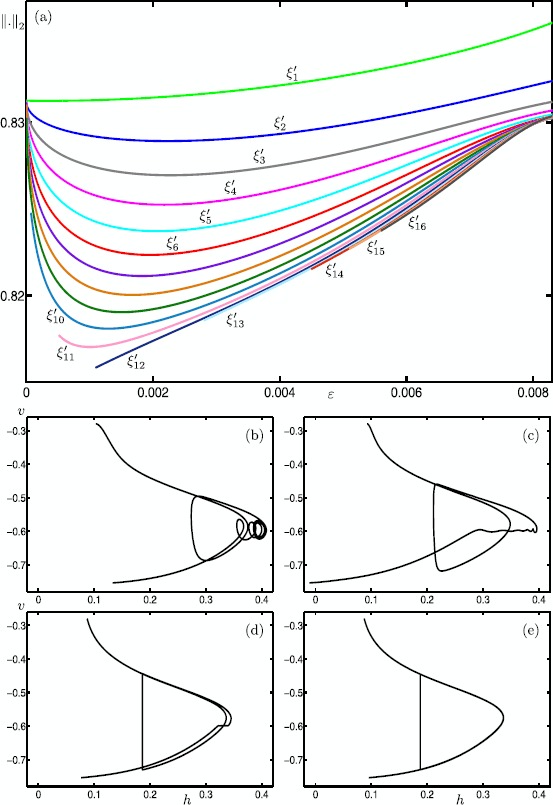
Many physiological phenomena have the property that some variables evolve much faster than others. For example, neuron models typically involve observable differences in time scales. The Hodgkin-Huxley model is well known for explaining the ionic mechanism that generates the action potential in the squid giant axon. Rubin and Wechselberger (Biol. Cybern. 97:5-32, 2007) nondimensionalized this model and obtained a singularly perturbed system with two fast, two slow variables, and an explicit time-scale ratio ε. The dynamics of this system are complex and feature periodic orbits with a series of action potentials separated by small-amplitude oscillations (SAOs); also referred to as mixed-mode oscillations (MMOs). The slow dynamics of this system are organized by two-dimensional locally invariant manifolds called slow manifolds which can be either attracting or of saddle type.In this paper, we introduce a general approach for computing two-dimensional saddle slow manifolds and their stable and unstable fast manifolds. We also develop a technique for detecting and continuing associated canard orbits, which arise from the interaction between attracting and saddle slow manifolds, and provide a mechanism for the organization of SAOs in [Formula: see text]. We first test our approach with an extended four-dimensional normal form of a folded node. Our results demonstrate that our computations give reliable approximations of slow manifolds and canard orbits of this model. Our computational approach is then utilized to investigate the role of saddle slow manifolds and associated canard orbits of the full Hodgkin-Huxley model in organizing MMOs and determining the firing rates of action potentials. For ε sufficiently large, canard orbits are arranged in pairs of twin canard orbits with the same number of SAOs. We illustrate how twin canard orbits partition the attracting slow manifold into a number of ribbons that play the role of sectors of rotations. The upshot is that we are able to unravel the geometry of slow manifolds and associated canard orbits without the need to reduce the model.
某些变量的演化速度比其他变量快得多。例如,神经元模型通常涉及时间尺度上的可观测差异。霍奇金 - 赫胥黎模型以解释鱿鱼巨轴突中产生动作电位的离子机制而闻名。鲁宾和韦chselberger(《生物控制论》97:5 - 32,2007)对该模型进行了无量纲化处理,得到了一个具有两个快速变量、两个慢速变量以及明确时间尺度比ε的奇异摄动系统。该系统的动力学很复杂,具有一系列由小振幅振荡(SAO)分隔的动作电位的周期轨道;也被称为混合模式振荡(MMO)。该系统的慢动力学由称为慢流形的二维局部不变流形组织,慢流形可以是吸引型或鞍型。在本文中,我们介绍了一种计算二维鞍型慢流形及其稳定和不稳定快流形的通用方法。我们还开发了一种检测和延续相关鸭轨道的技术,鸭轨道由吸引型和鞍型慢流形之间的相互作用产生,并为[公式:见原文]中SAO的组织提供了一种机制。我们首先用折叠节点的扩展四维范式测试我们的方法。我们的结果表明,我们的计算给出了该模型慢流形和鸭轨道的可靠近似。然后,我们利用我们的计算方法来研究完整霍奇金 - 赫胥黎模型的鞍型慢流形和相关鸭轨道在组织MMO和确定动作电位发放率中的作用。对于足够大的ε,鸭轨道成对排列成具有相同数量SAO的双鸭轨道。我们说明了双鸭轨道如何将吸引型慢流形划分为多个起旋转扇区作用的条带。结果是,我们能够在无需简化模型的情况下揭示慢流形和相关鸭轨道的几何结构。