The Xiphophorus Genetic Stock Center, Molecular Biosciences Research Group, Department of Chemistry and Biochemistry, Texas State University, 419 Centennial Hall, 601 University Drive, San Marcos, TX, 78666, USA.
BMC Genomics. 2018 May 10;19(1):355. doi: 10.1186/s12864-018-4735-5.
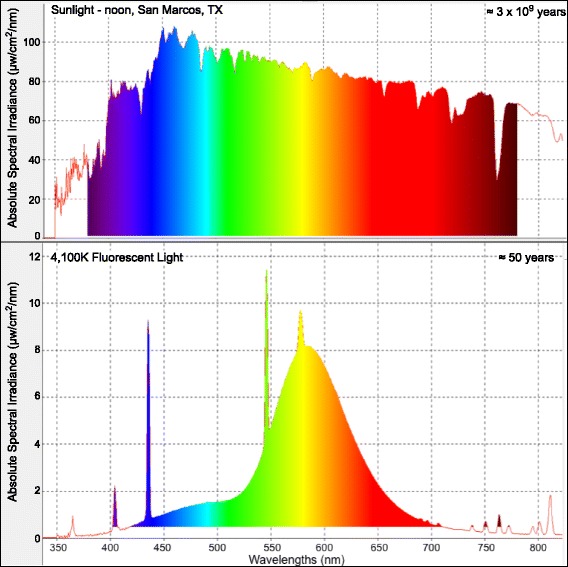
Evolution occurred exclusively under the full spectrum of sunlight. Conscription of narrow regions of the solar spectrum by specific photoreceptors suggests a common strategy for regulation of genetic pathways. Fluorescent light (FL) does not possess the complexity of the solar spectrum and has only been in service for about 60 years. If vertebrates evolved specific genetic responses regulated by light wavelengths representing the entire solar spectrum, there may be genetic consequences to reducing the spectral complexity of light.
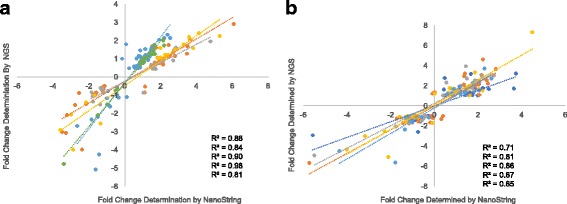
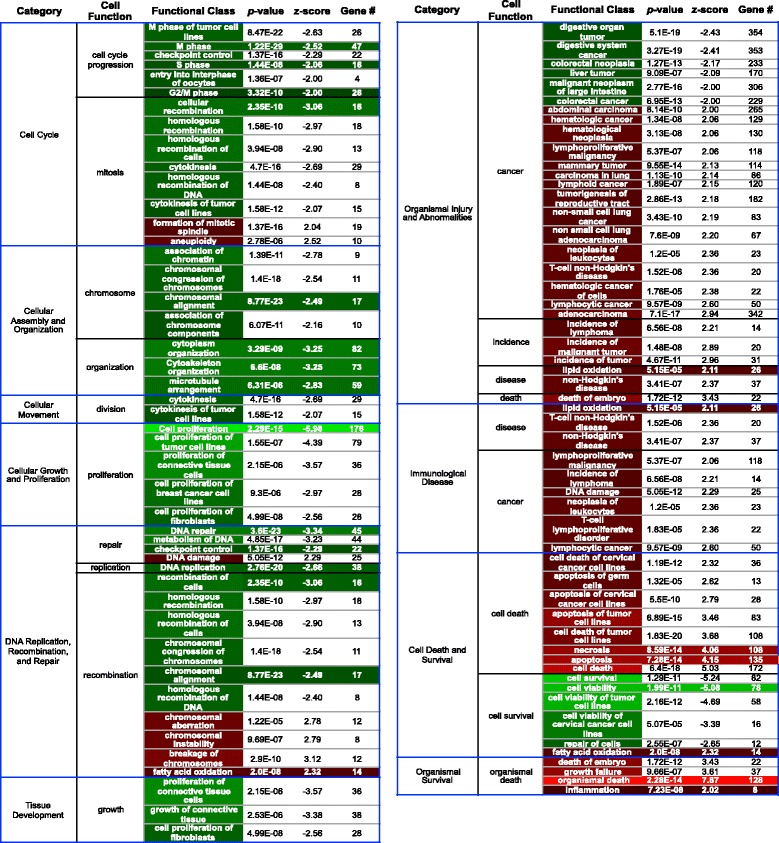
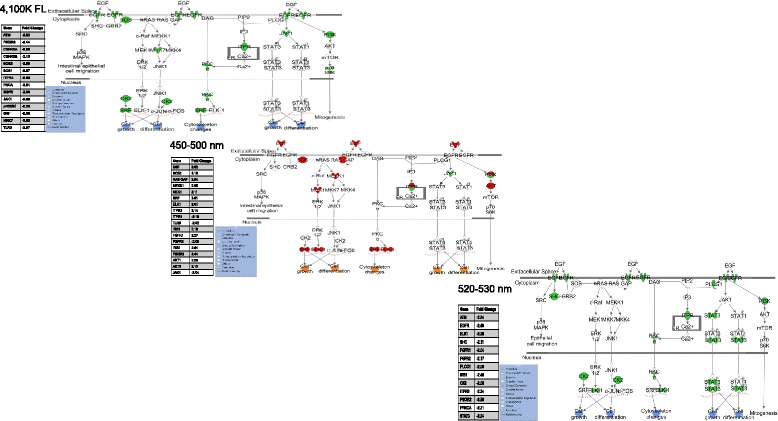
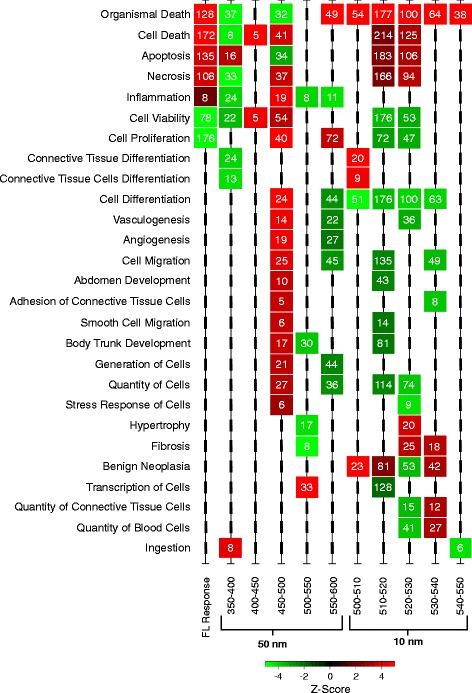
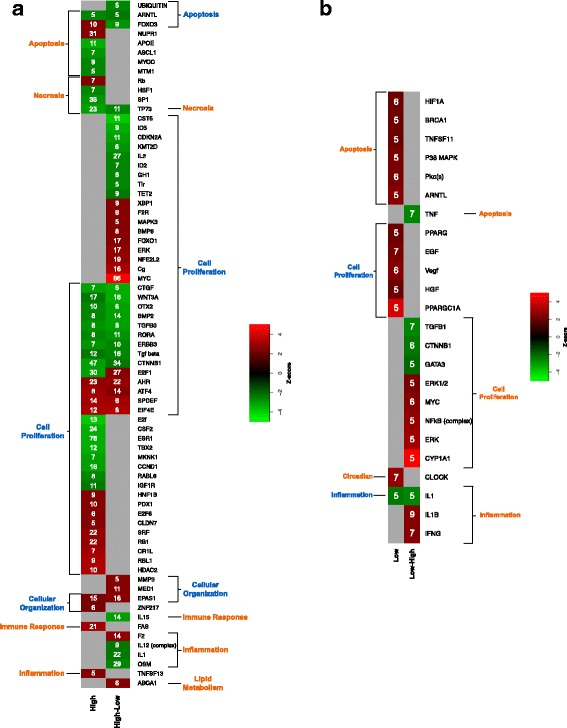
We utilized RNA-Seq to assess changes in the transcriptional profiles of Xiphophorus maculatus skin after exposure to FL ("cool white"), or narrow wavelength regions of light between 350 and 600 nm (i.e., 50 nm or 10 nm regions, herein termed "wavebands"). Exposure to each 50 nm waveband identified sets of genes representing discrete pathways that showed waveband specific transcriptional modulation. For example, 350-400 or 450-500 nm waveband exposures resulted in opposite regulation of gene sets marking necrosis and apoptosis (i.e., 350-400 nm; necrosis suppression, apoptosis activation, while 450-500 nm; apoptosis suppression, necrosis activation). Further investigation of specific transcriptional modulation employing successive 10 nm waveband exposures between 500 and 550 nm showed; (a) greater numbers of genes may be transcriptionally modulated after 10 nm exposures, than observed for 50 nm or FL exposures, (b) the 10 nm wavebands induced gene sets showing greater functional specificity than 50 nm or FL exposures, and (c) the genetic effects of FL are primarily due to 30 nm between 500 and 530 nm. Interestingly, many genetic pathways exhibited completely opposite transcriptional effects after different waveband exposures. For example, the epidermal growth factor (EGF) pathway exhibits transcriptional suppression after FL exposure, becomes highly active after 450-500 nm waveband exposure, and again, exhibits strong transcriptional suppression after exposure to the 520-530 nm waveband.
Collectively, these results suggest one may manipulate transcription of specific genetic pathways in skin by exposure of the intact animal to specific wavebands of light. In addition, we identify genes transcriptionally modulated in a predictable manner by specific waveband exposures. Such genes, and their regulatory elements, may represent valuable tools for genetic engineering and gene therapy protocols.
进化完全是在阳光的全光谱下发生的。特定光感受器对太阳光谱的特定窄区域的吸收表明,遗传途径的调控存在一种共同的策略。荧光灯(FL)不具备太阳光谱的复杂性,并且仅使用了大约 60 年。如果脊椎动物进化出了受代表整个太阳光谱的光波长调节的特定遗传反应,那么减少光的光谱复杂性可能会产生遗传后果。
我们利用 RNA-Seq 来评估暴露于 FL(“冷白”)或 350 至 600nm 之间的窄波长区域的光(即 50nm 或 10nm 区域,在此称为“波段”)后,新月锦鱼皮肤的转录谱的变化。暴露于每个 50nm 波段会确定代表离散途径的基因集,这些途径表现出波段特异性的转录调节。例如,350-400nm 或 450-500nm 波段的暴露导致标记坏死和凋亡的基因集的相反调节(即 350-400nm;坏死抑制,凋亡激活,而 450-500nm;凋亡抑制,坏死激活)。通过在 500 至 550nm 之间使用连续的 10nm 波段进行特定转录调节的进一步研究表明;(a)与 50nm 或 FL 暴露相比,10nm 暴露后可能会转录调节更多数量的基因,(b)10nm 波段诱导的基因集比 50nm 或 FL 暴露表现出更大的功能特异性,并且(c)FL 的遗传效应主要归因于 500 至 530nm 之间的 30nm。有趣的是,许多遗传途径在不同的波段暴露后表现出完全相反的转录效应。例如,表皮生长因子(EGF)途径在 FL 暴露后表现出转录抑制,在 450-500nm 波段暴露后变得高度活跃,并且再次在暴露于 520-530nm 波段后表现出强烈的转录抑制。
总的来说,这些结果表明,通过将完整的动物暴露于特定的光波段,可以操纵皮肤中特定遗传途径的转录。此外,我们确定了以可预测的方式受特定波段暴露调节的转录基因。这些基因及其调节元件可能代表遗传工程和基因治疗方案的有价值的工具。