Department of Chemistry, Massachusetts Institute of Technology, Cambridge, United States.
Fred Hutchinson Cancer Research Center, Seattle, United States.
Elife. 2018 Sep 6;7:e38795. doi: 10.7554/eLife.38795.
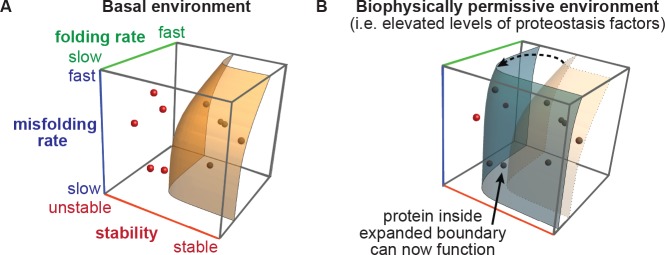
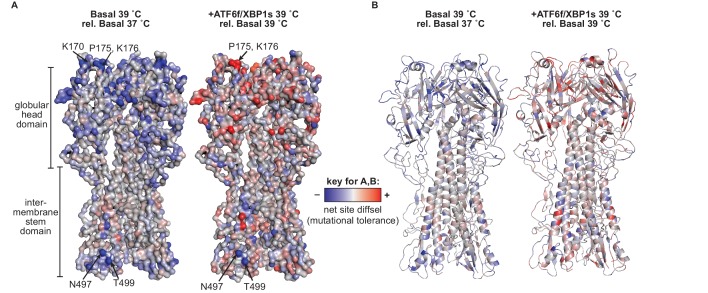
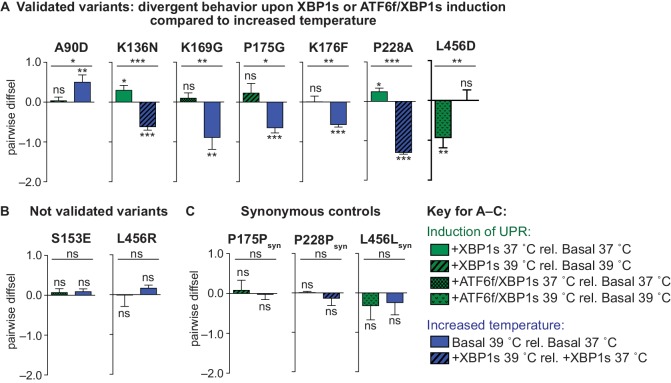
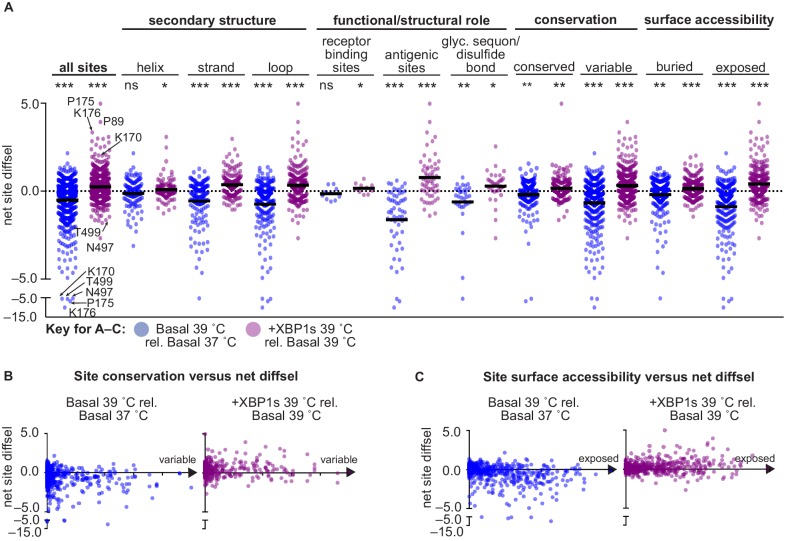
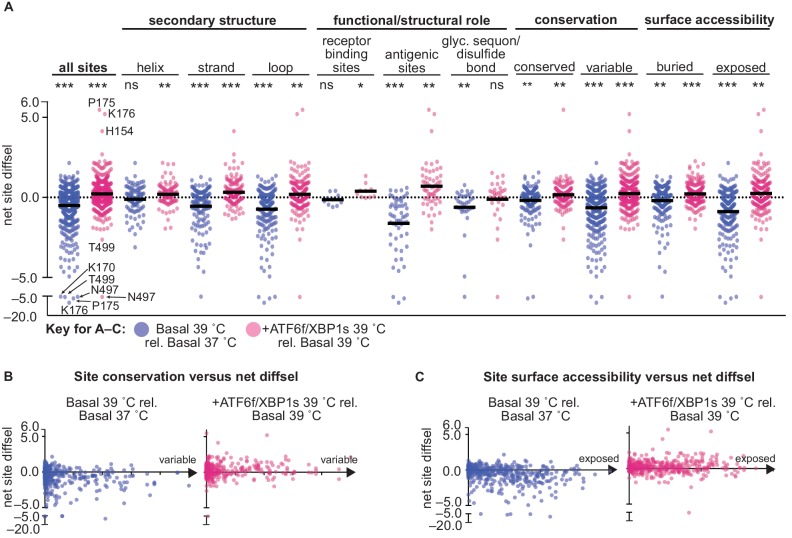
We systematically and quantitatively evaluate whether endoplasmic reticulum (ER) proteostasis factors impact the mutational tolerance of secretory pathway proteins. We focus on influenza hemaggluttinin (HA), a viral membrane protein that folds in the host's ER via a complex pathway. By integrating chemical methods to modulate ER proteostasis with deep mutational scanning to assess mutational tolerance, we discover that upregulation of ER proteostasis factors broadly enhances HA mutational tolerance across diverse structural elements. Remarkably, this proteostasis network-enhanced mutational tolerance occurs at the same sites where mutational tolerance is most reduced by propagation at fever-like temperature. These findings have important implications for influenza evolution, because influenza immune escape is contingent on HA possessing sufficient mutational tolerance to evade antibodies while maintaining the capacity to fold and function. More broadly, this work provides the first experimental evidence that ER proteostasis mechanisms define the mutational tolerance and, therefore, the evolution of secretory pathway proteins.
我们系统地和定量地评估内质网(ER)蛋白稳态因子是否影响分泌途径蛋白的突变耐受性。我们专注于流感血凝素(HA),一种通过复杂途径在宿主 ER 中折叠的病毒膜蛋白。通过整合化学方法来调节 ER 蛋白稳态和深度突变扫描来评估突变耐受性,我们发现 ER 蛋白稳态因子的上调广泛增强了 HA 的突变耐受性,跨越了多种结构元件。值得注意的是,这种蛋白质稳态网络增强的突变耐受性发生在同样的位点,在这些位点,通过发热样温度的传播最能降低突变耐受性。这些发现对流感进化具有重要意义,因为流感免疫逃逸取决于 HA 具有足够的突变耐受性,以逃避抗体,同时保持折叠和功能的能力。更广泛地说,这项工作提供了第一个实验证据,证明 ER 蛋白质稳态机制定义了突变耐受性,因此也定义了分泌途径蛋白的进化。