Department of Molecular, Cell and Developmental Biology, University of California, Los Angeles, Los Angeles, United States.
The Sainsbury Laboratory, University of Cambridge, Cambridge, United Kingdom.
Elife. 2018 Sep 18;7:e38161. doi: 10.7554/eLife.38161.
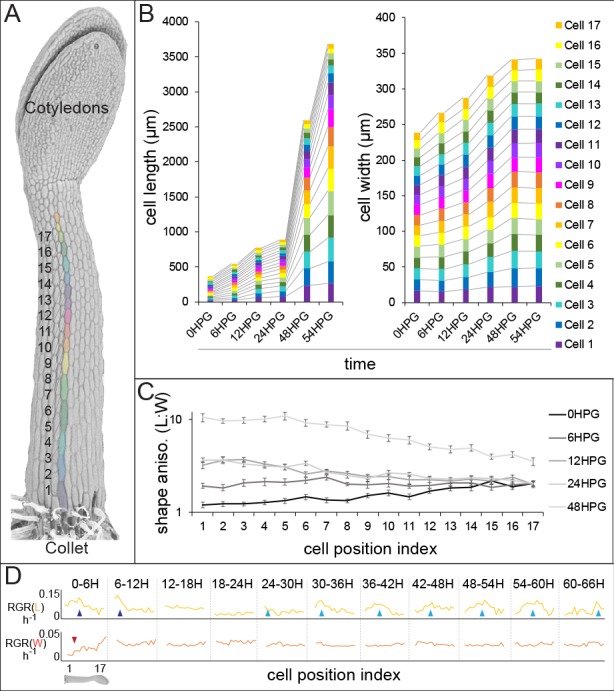
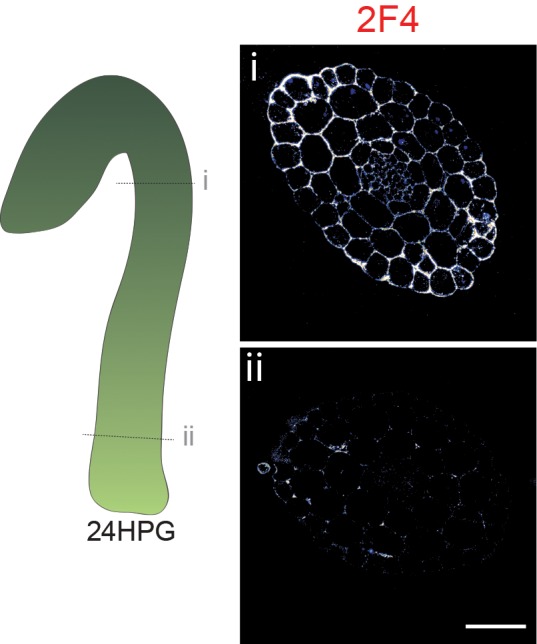
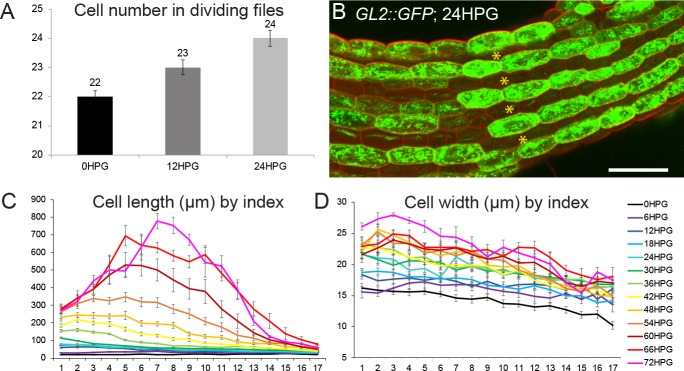
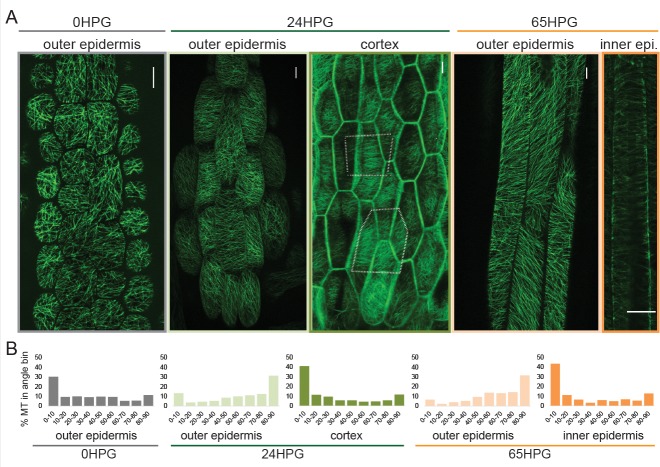
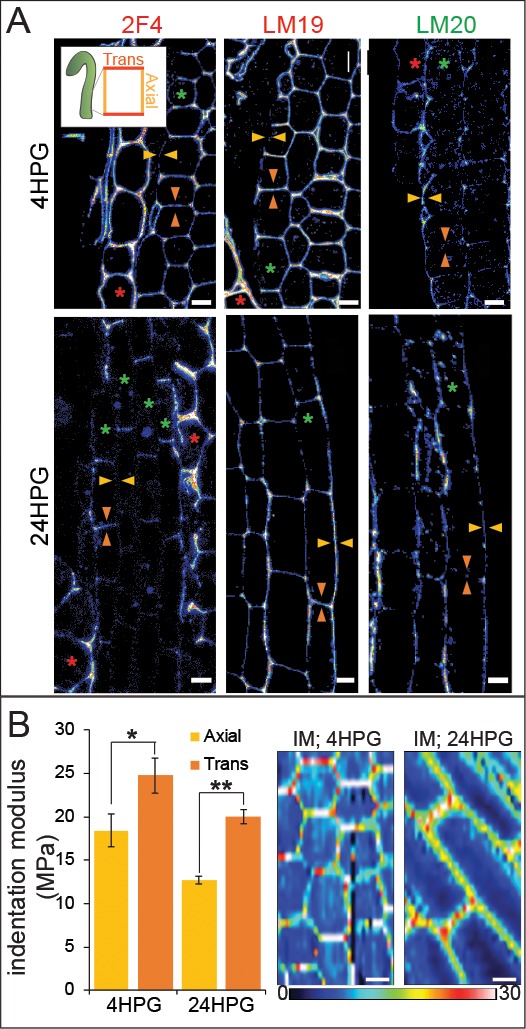
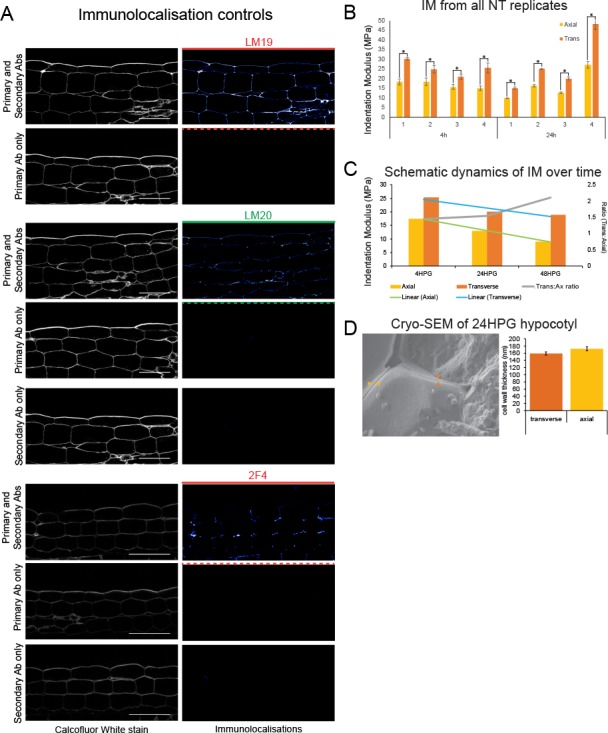
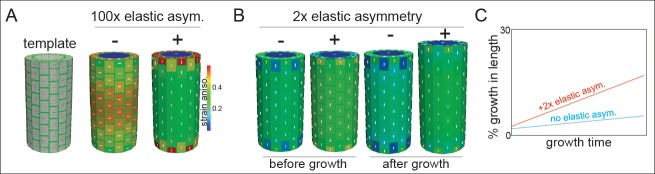
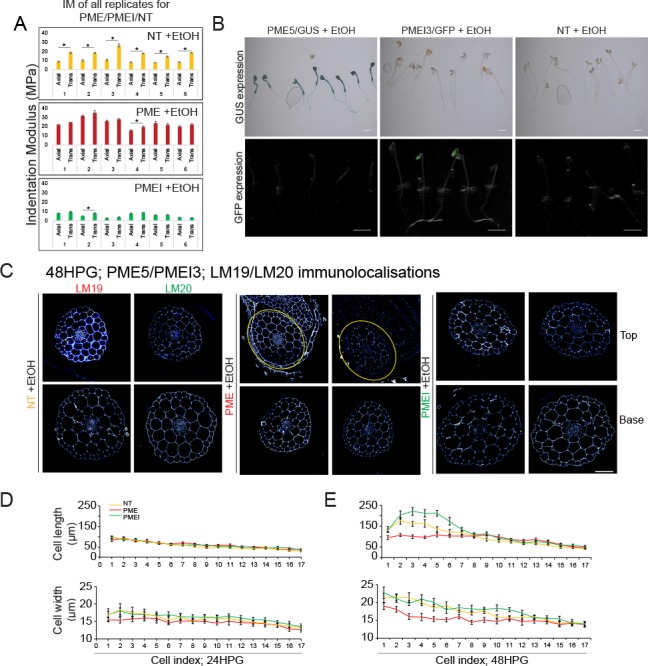
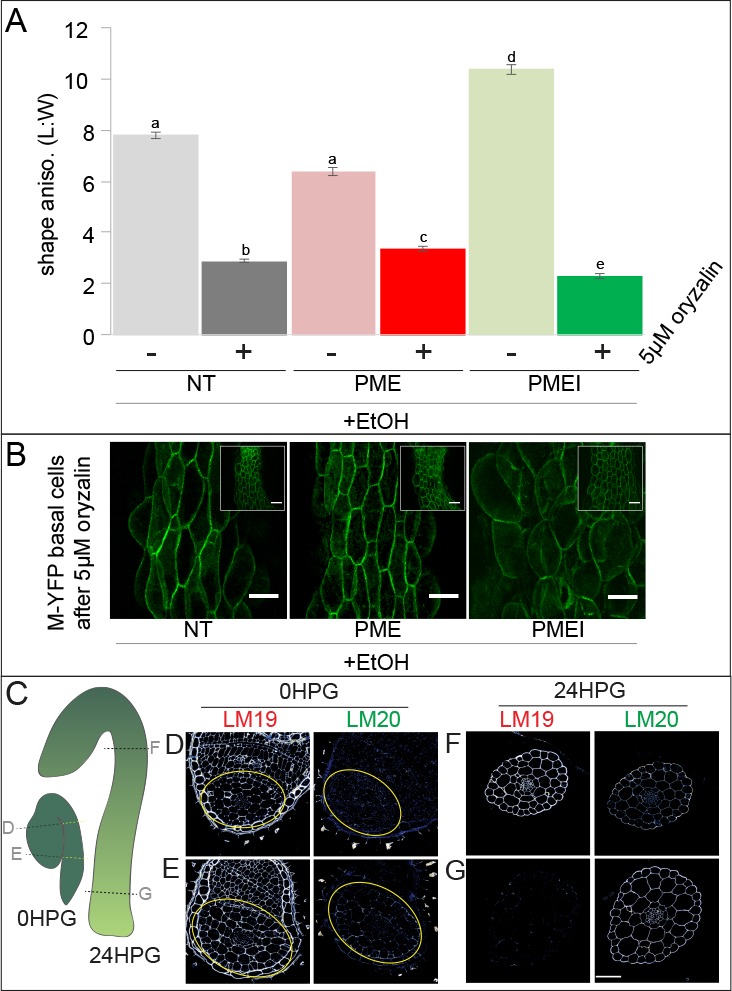
Fast directional growth is a necessity for the young seedling; after germination, it needs to quickly penetrate the soil to begin its autotrophic life. In most dicot plants, this rapid escape is due to the anisotropic elongation of the hypocotyl, the columnar organ between the root and the shoot meristems. Anisotropic growth is common in plant organs and is canonically attributed to cell wall anisotropy produced by oriented cellulose fibers. Recently, a mechanism based on asymmetric pectin-based cell wall elasticity has been proposed. Here we present a harmonizing model for anisotropic growth control in the dark-grown hypocotyl: basic anisotropic information is provided by cellulose orientation) and additive anisotropic information is provided by pectin-based elastic asymmetry in the epidermis. We quantitatively show that hypocotyl elongation is anisotropic starting at germination. We present experimental evidence for pectin biochemical differences and wall mechanics providing important growth regulation in the hypocotyl. Lastly, our in silico modelling experiments indicate an additive collaboration between pectin biochemistry and cellulose orientation in promoting anisotropic growth.
快速的方向性生长是幼苗的必要条件;发芽后,它需要迅速穿透土壤,开始自养生活。在大多数双子叶植物中,这种快速的逃逸是由于下胚轴的各向异性伸长引起的,下胚轴是根和茎分生组织之间的柱状器官。各向异性生长在植物器官中很常见,通常归因于取向纤维素纤维产生的细胞壁各向异性。最近,提出了一种基于不对称果胶细胞壁弹性的机制。在这里,我们提出了一个协调模型,用于黑暗生长的下胚轴的各向异性生长控制:基本的各向异性信息由纤维素取向提供,附加的各向异性信息由表皮中的基于果胶的弹性不对称性提供。我们定量地表明,从发芽开始,下胚轴的伸长就是各向异性的。我们提供了果胶生化差异和细胞壁力学的实验证据,这些证据为下胚轴的重要生长调节提供了证据。最后,我们的计算机模拟实验表明,果胶生化和纤维素取向在促进各向异性生长方面具有附加的协同作用。