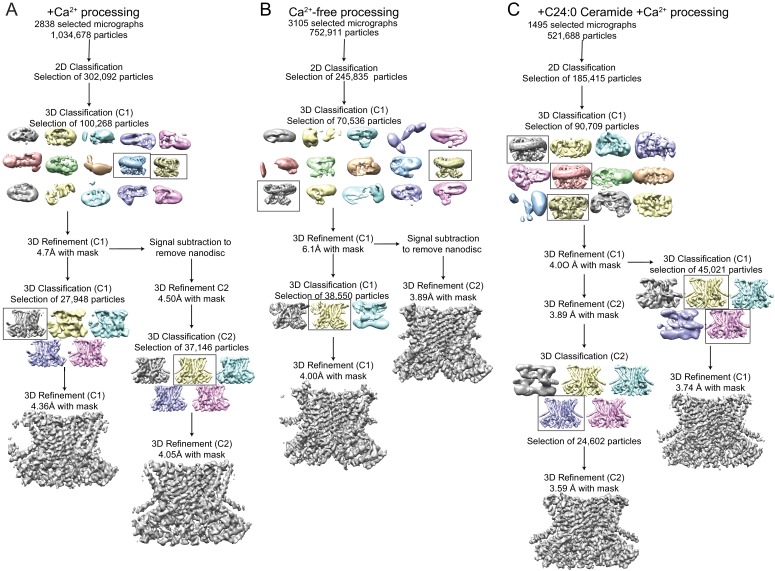
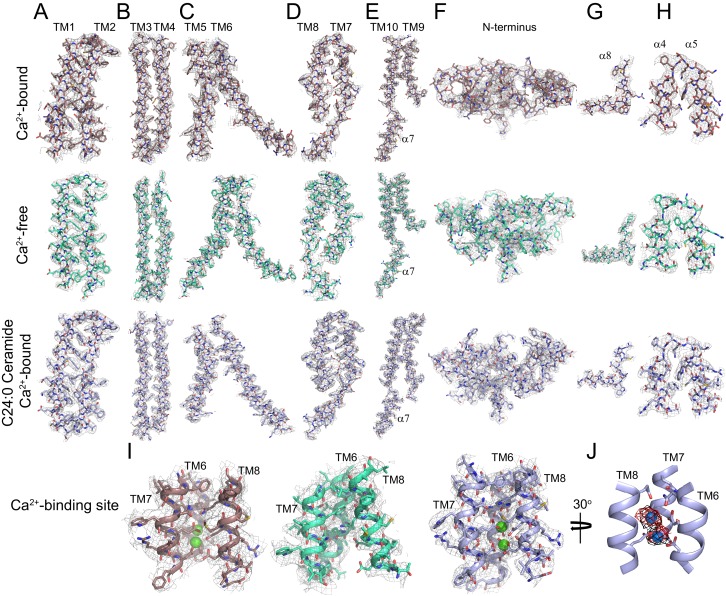
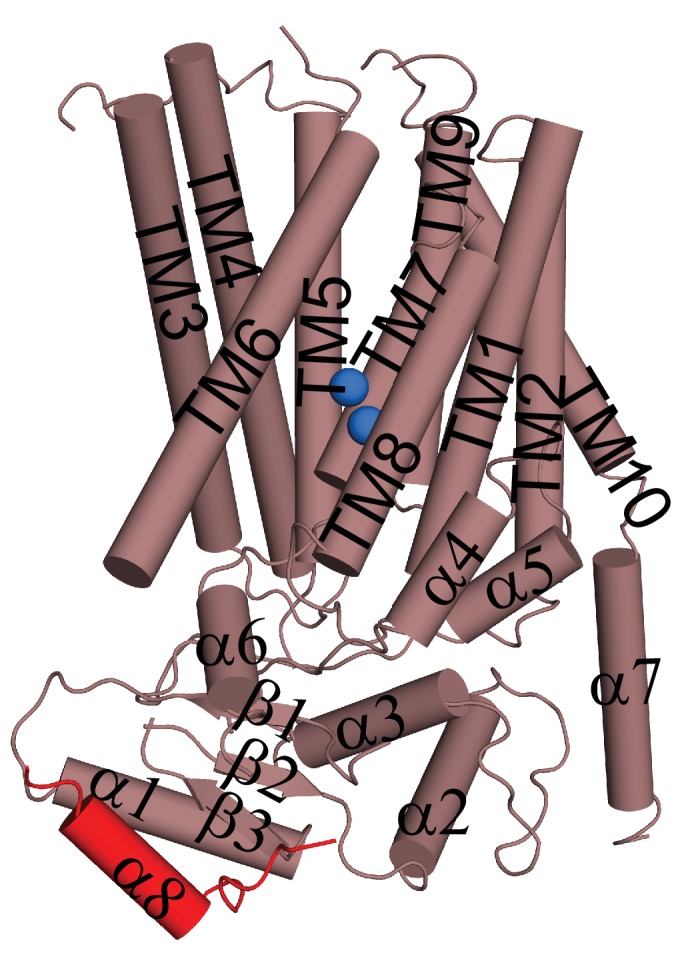
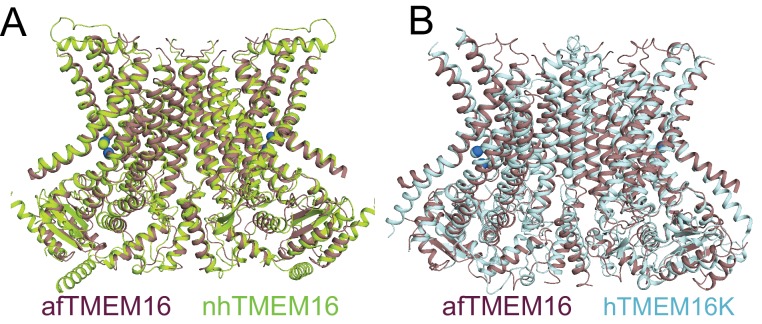
Department of Biochemistry, Weill Cornell Medical College, New York, United States.
Department of Anesthesiology, Weill Cornell Medical College, New York, United States.
Elife. 2019 Jan 16;8:e43229. doi: 10.7554/eLife.43229.
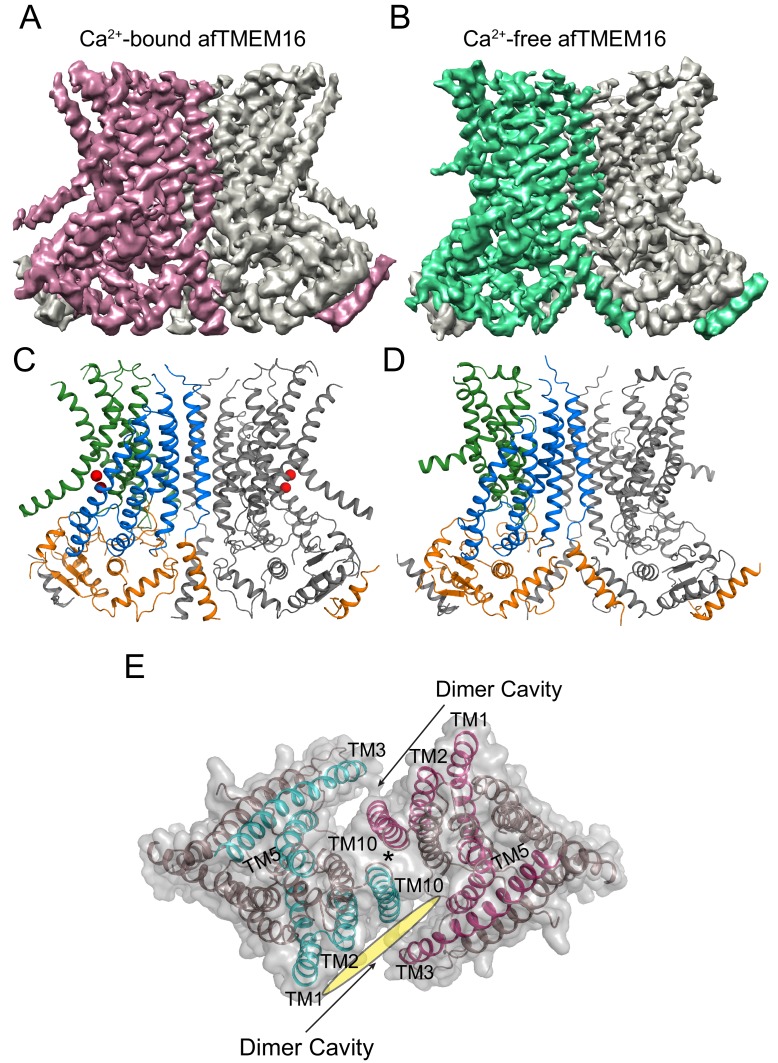
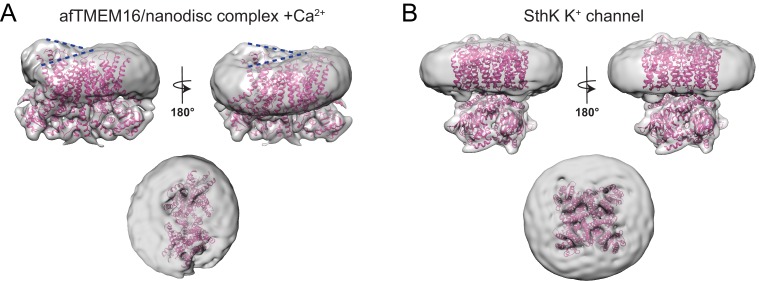
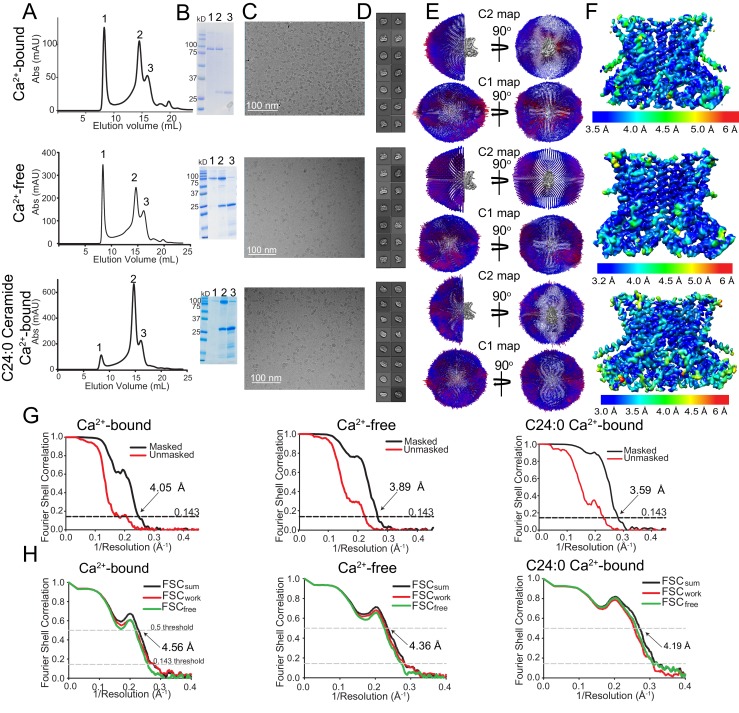
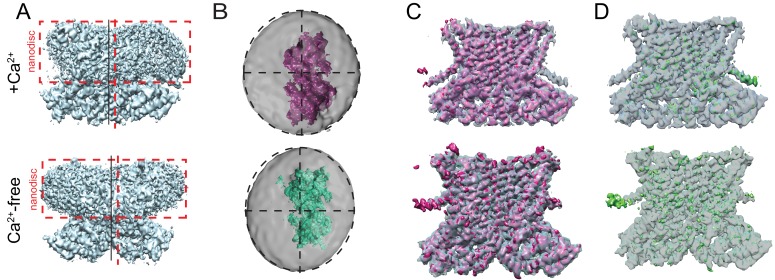
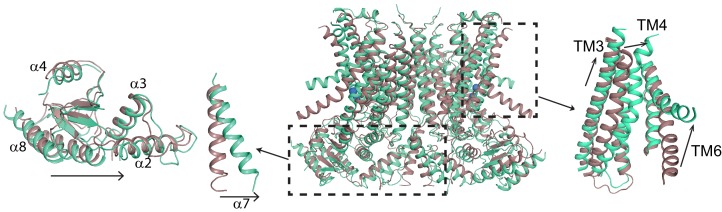
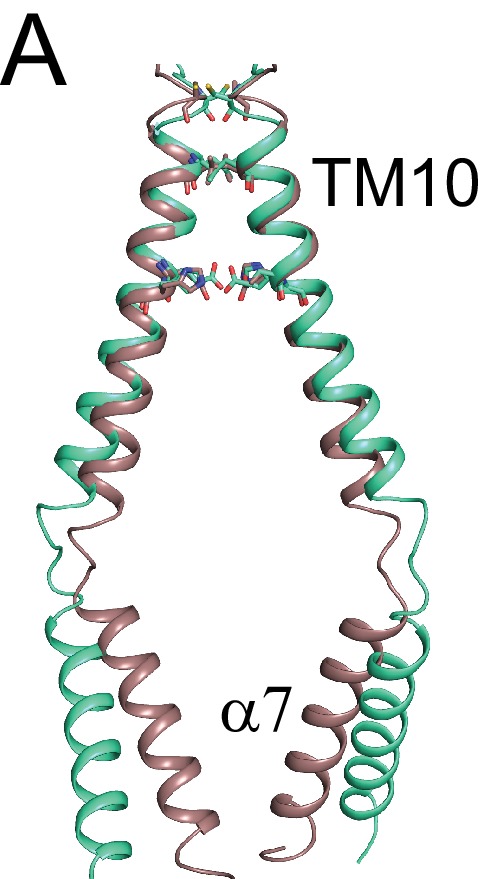
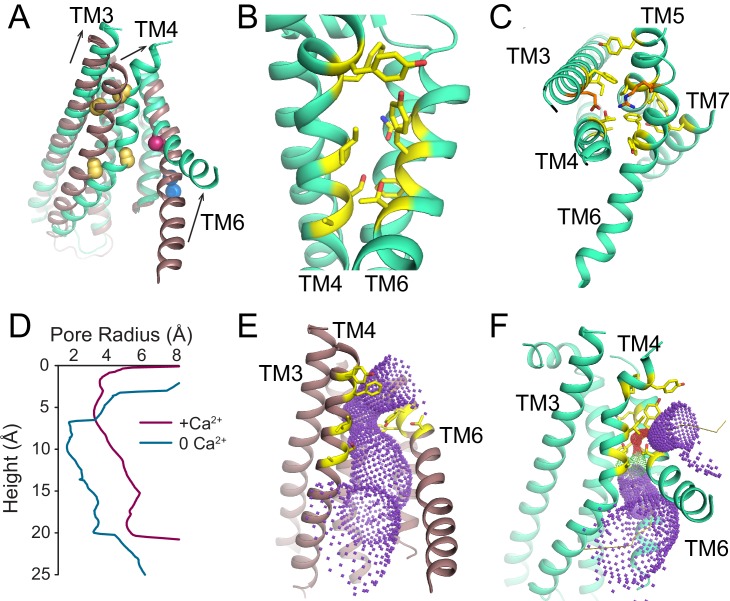
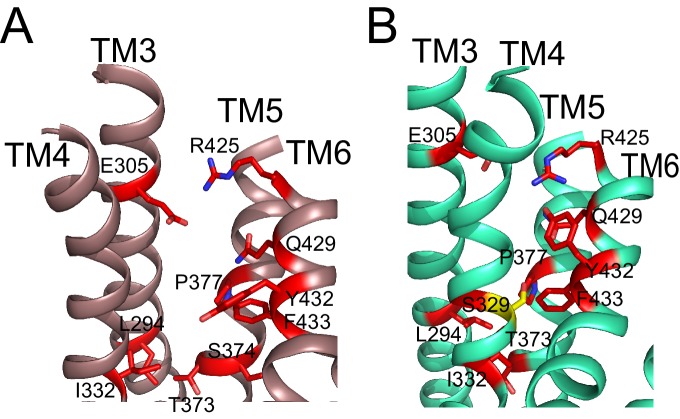
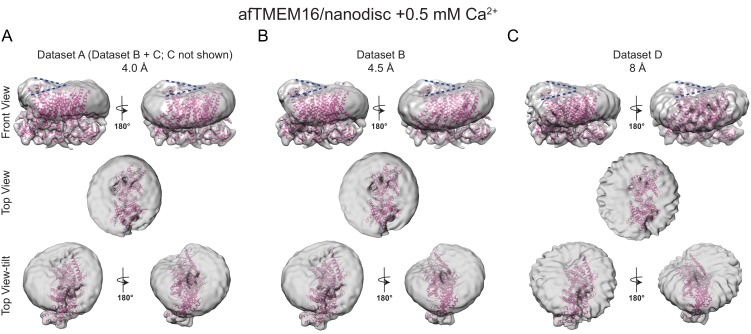
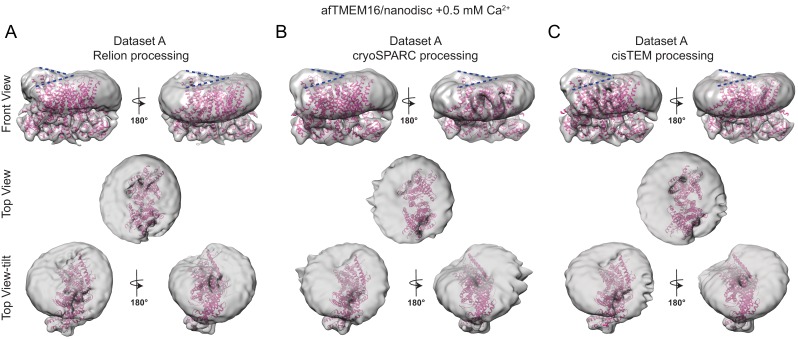
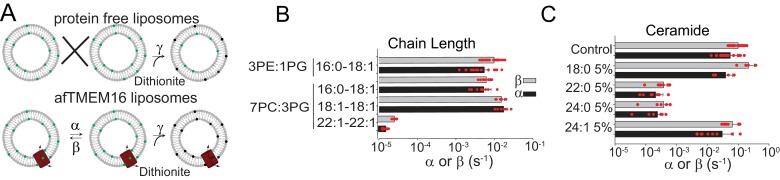
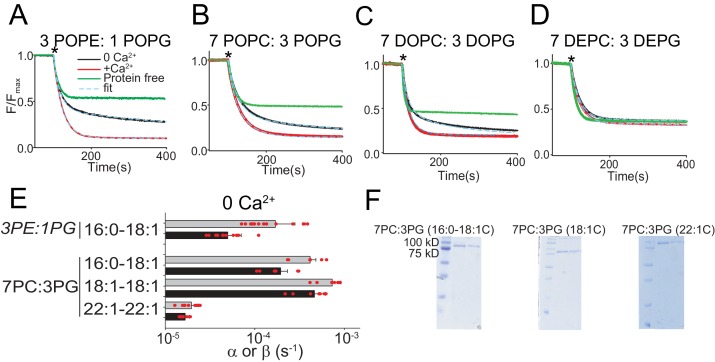
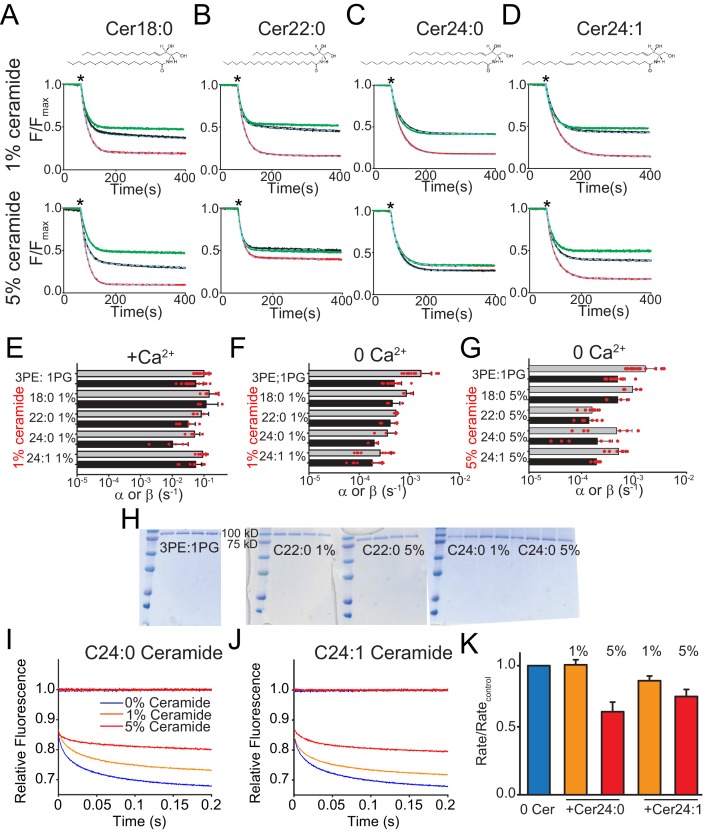
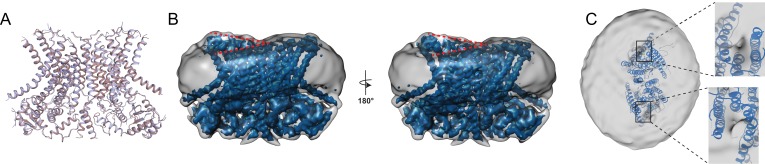
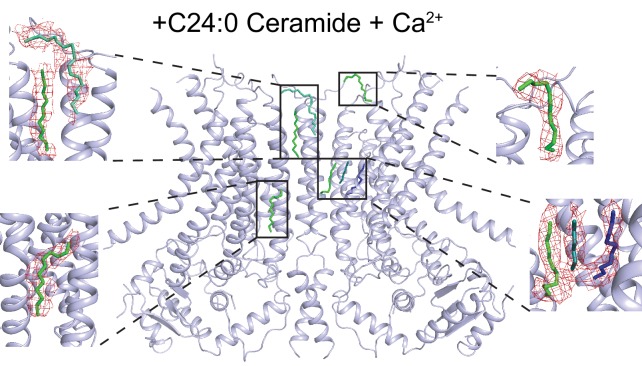
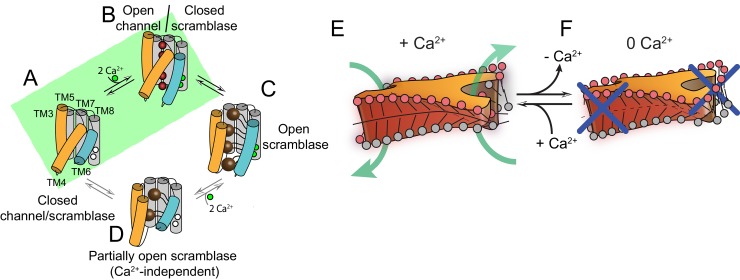
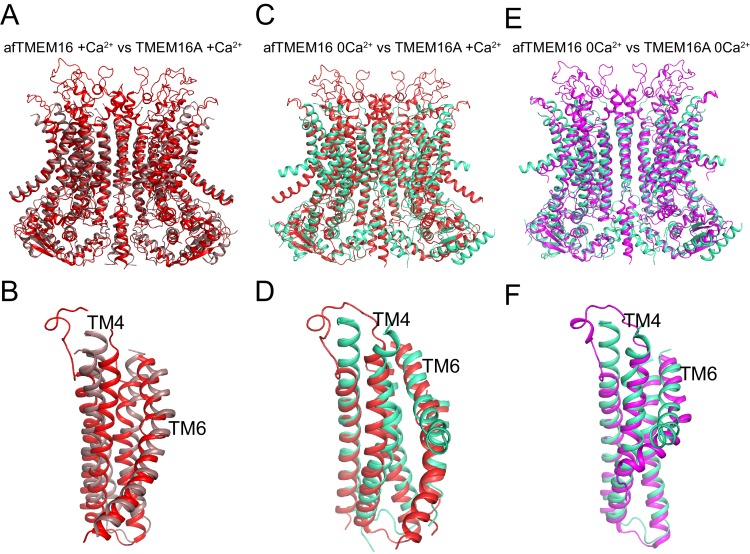
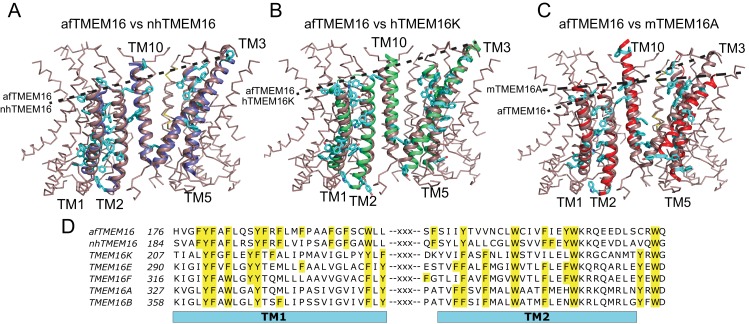
The lipid distribution of plasma membranes of eukaryotic cells is asymmetric and phospholipid scramblases disrupt this asymmetry by mediating the rapid, nonselective transport of lipids down their concentration gradients. As a result, phosphatidylserine is exposed to the outer leaflet of membrane, an important step in extracellular signaling networks controlling processes such as apoptosis, blood coagulation, membrane fusion and repair. Several TMEM16 family members have been identified as Ca-activated scramblases, but the mechanisms underlying their Ca-dependent gating and their effects on the surrounding lipid bilayer remain poorly understood. Here, we describe three high-resolution cryo-electron microscopy structures of a fungal scramblase from , afTMEM16, reconstituted in lipid nanodiscs. These structures reveal that Ca-dependent activation of the scramblase entails global rearrangement of the transmembrane and cytosolic domains. These structures, together with functional experiments, suggest that activation of the protein thins the membrane near the transport pathway to facilitate rapid transbilayer lipid movement.
真核细胞质膜的脂质分布呈不对称性,而磷脂翻转酶通过介导脂质沿浓度梯度的快速非选择性运输来破坏这种不对称性。结果,磷脂酰丝氨酸暴露于膜的外叶,这是细胞外信号网络控制凋亡、血液凝固、膜融合和修复等过程的重要步骤。已经鉴定出几种 TMEM16 家族成员作为 Ca 激活的翻转酶,但它们的 Ca 依赖性门控机制及其对周围脂质双层的影响仍知之甚少。在这里,我们描述了真菌翻转酶 afTMEM16 在脂质纳米盘中重建的三个高分辨率冷冻电子显微镜结构。这些结构表明,Ca 依赖性翻转酶的激活需要跨膜和胞质结构域的全局重排。这些结构与功能实验一起表明,蛋白质的激活会使靠近运输途径的膜变薄,以促进快速的跨膜脂质运动。