Interdisciplinary Biological Sciences Program, Northwestern University, Evanston, United States.
Department of Molecular Biosciences, Northwestern University, Evanston, United States.
Elife. 2019 Apr 8;8:e40260. doi: 10.7554/eLife.40260.
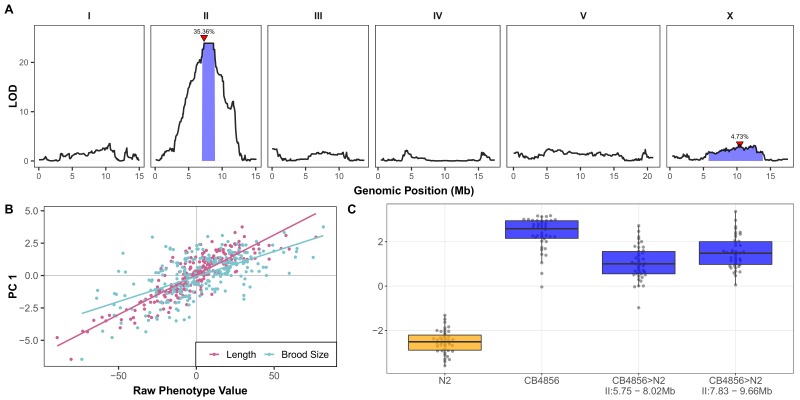
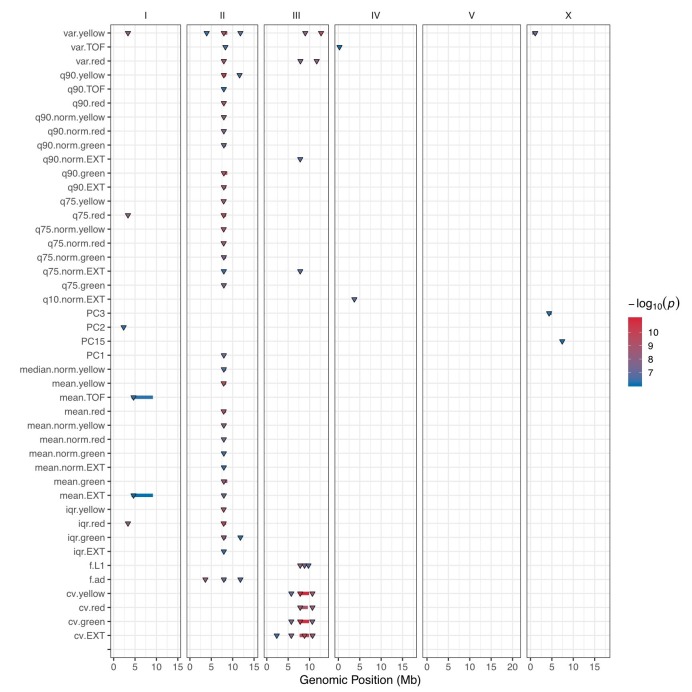
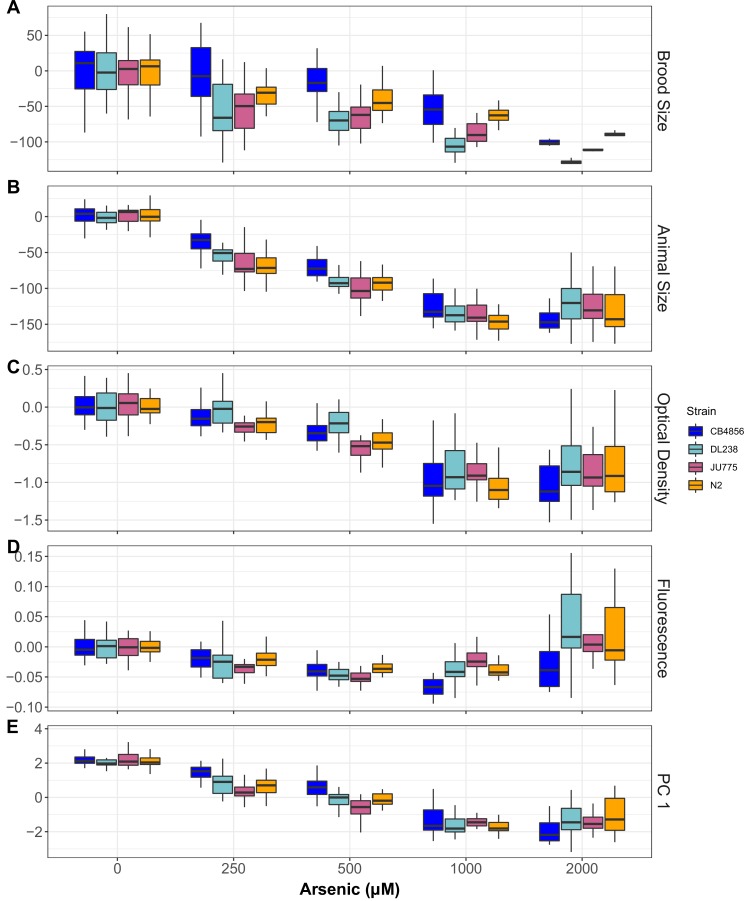
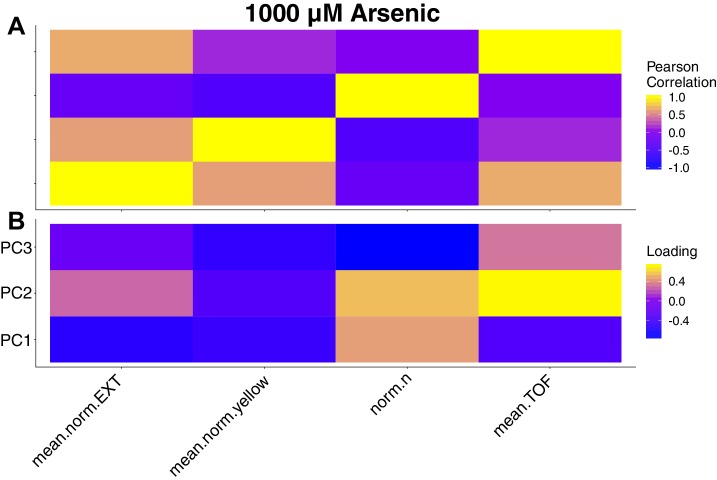
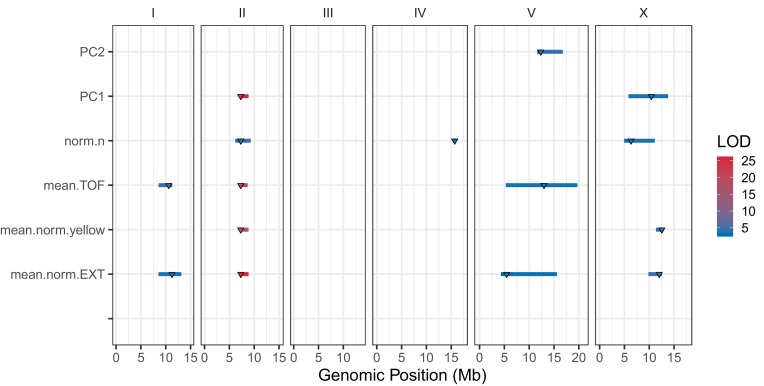
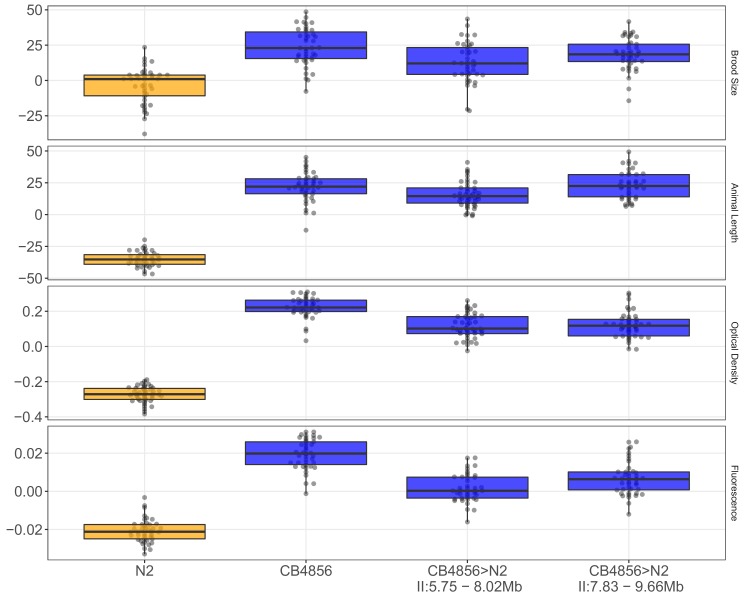
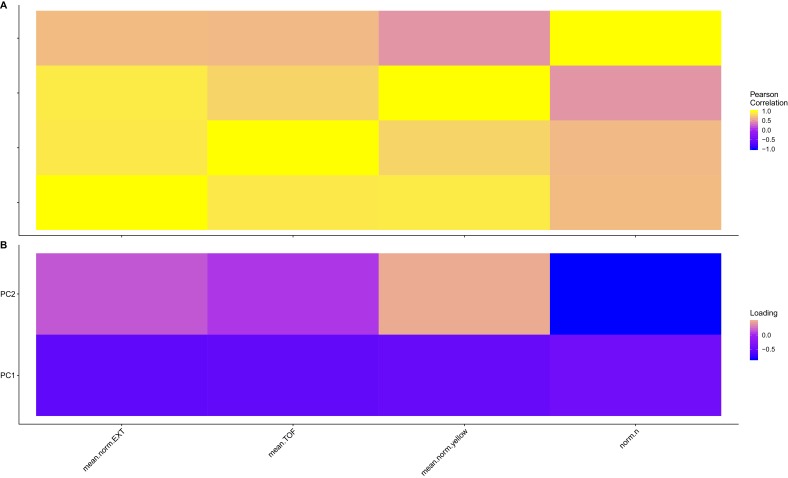
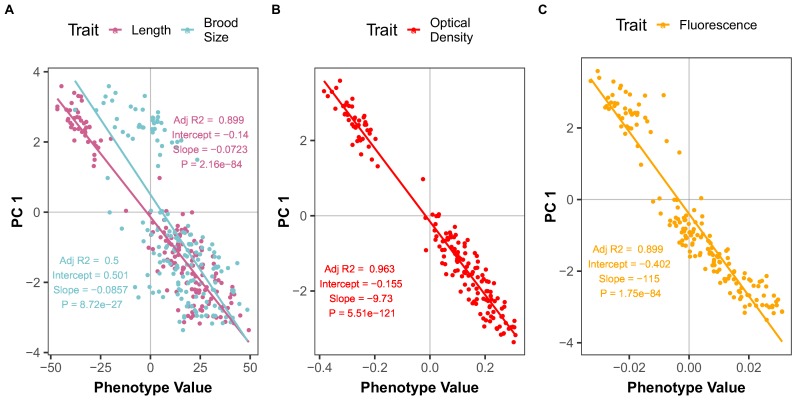
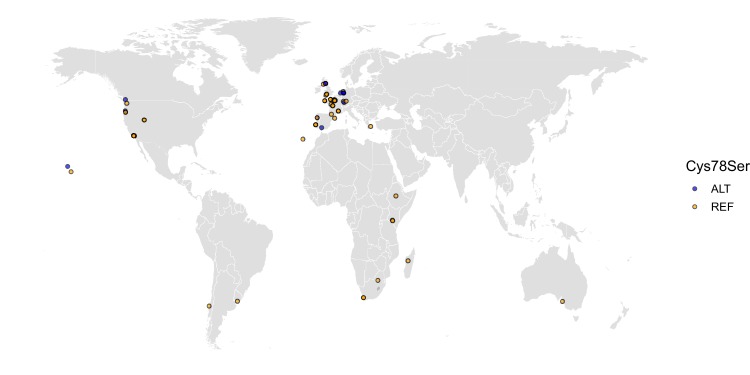
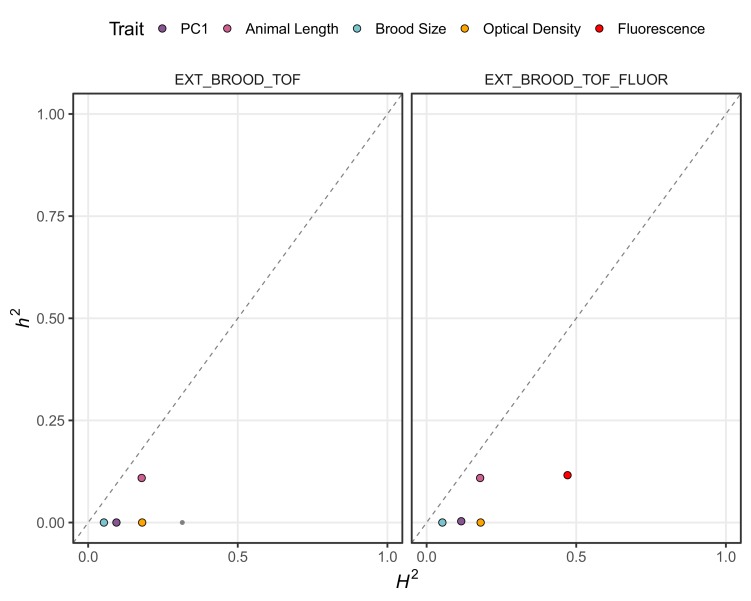
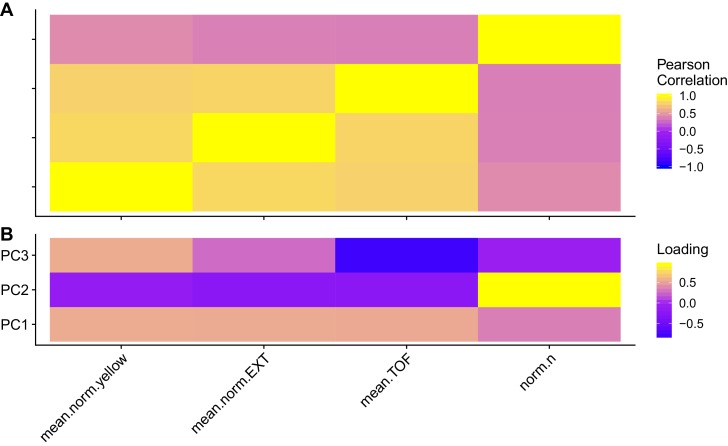
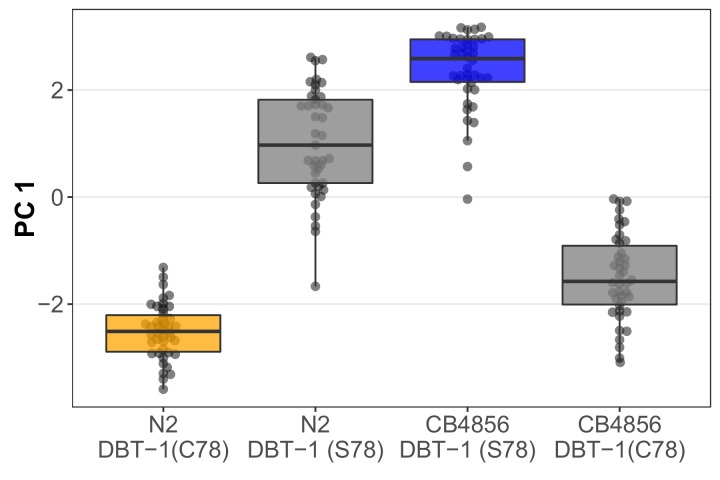
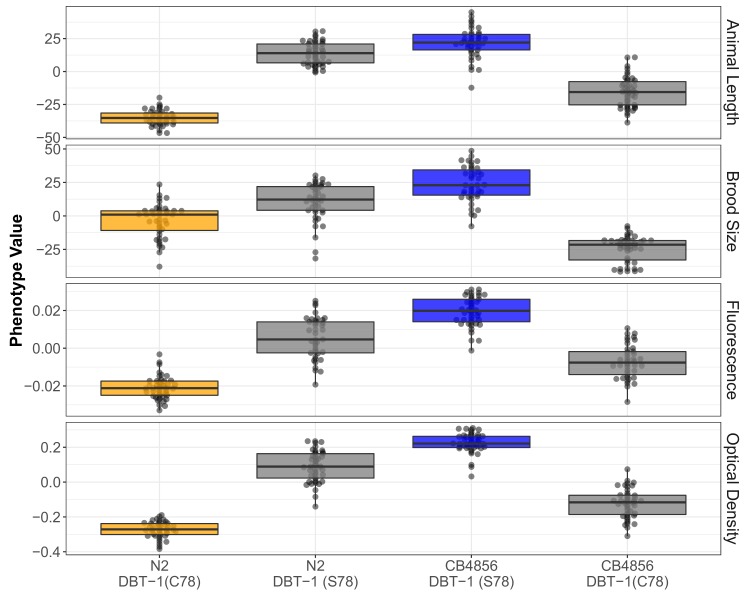
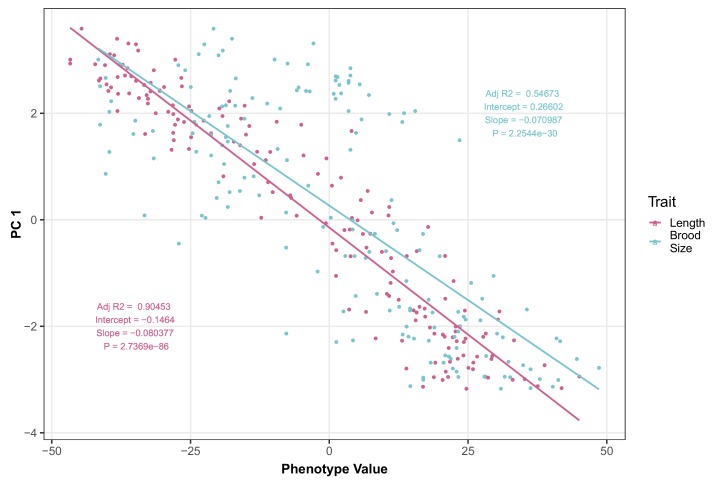
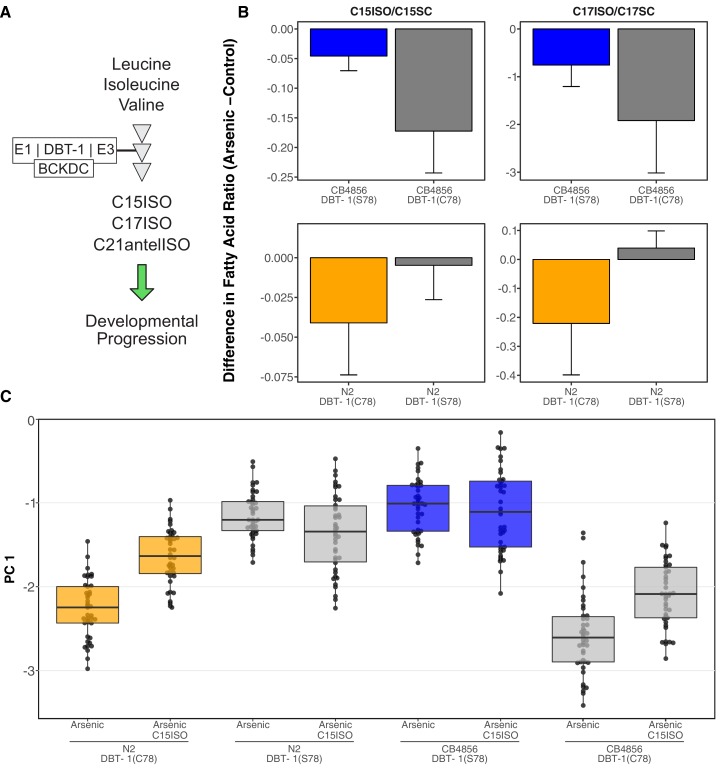
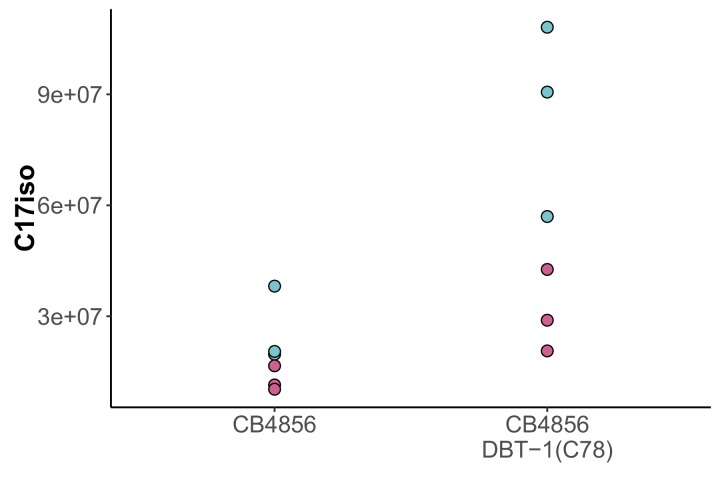
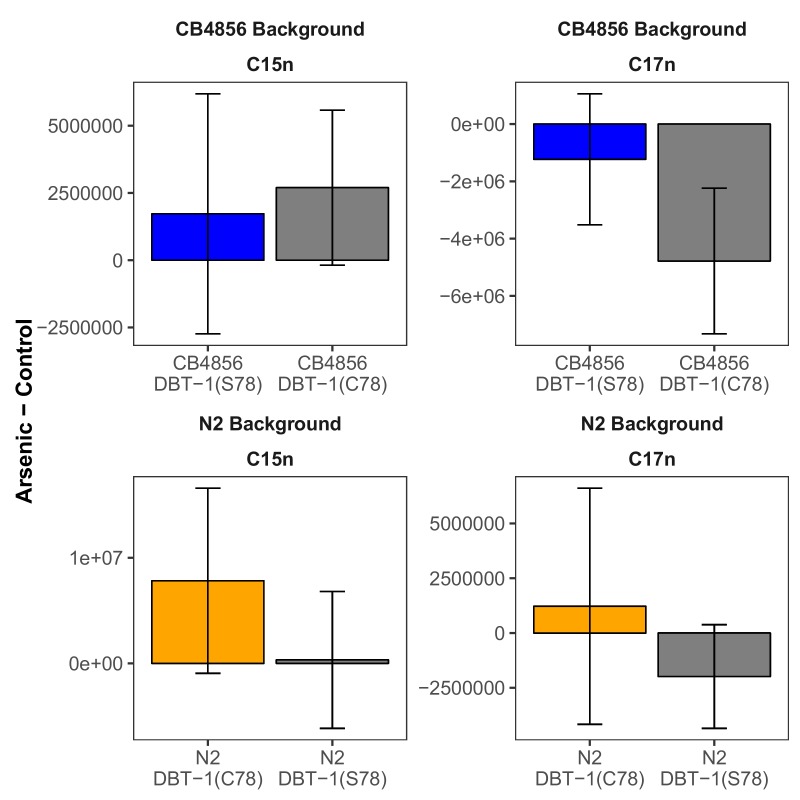
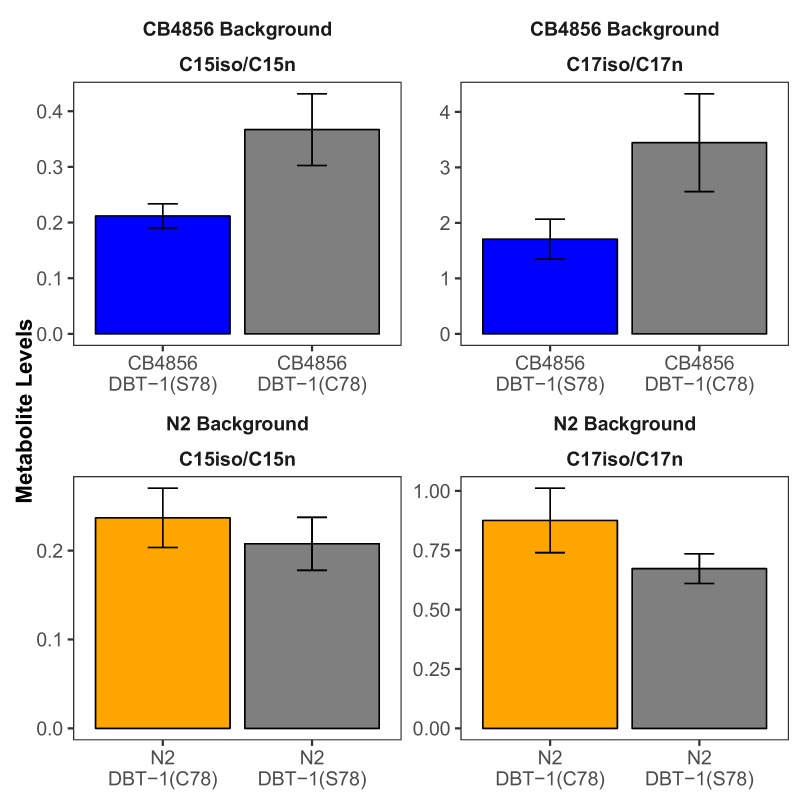
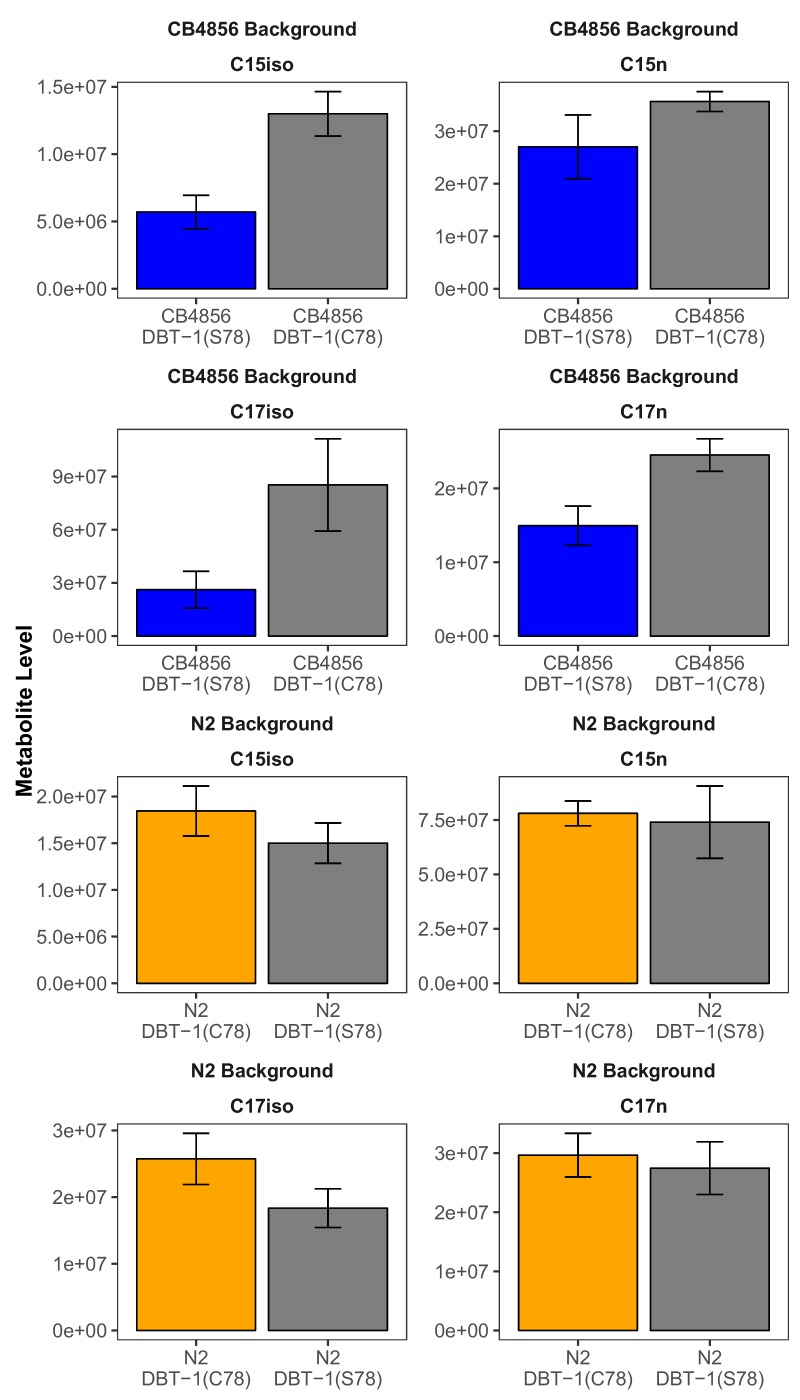
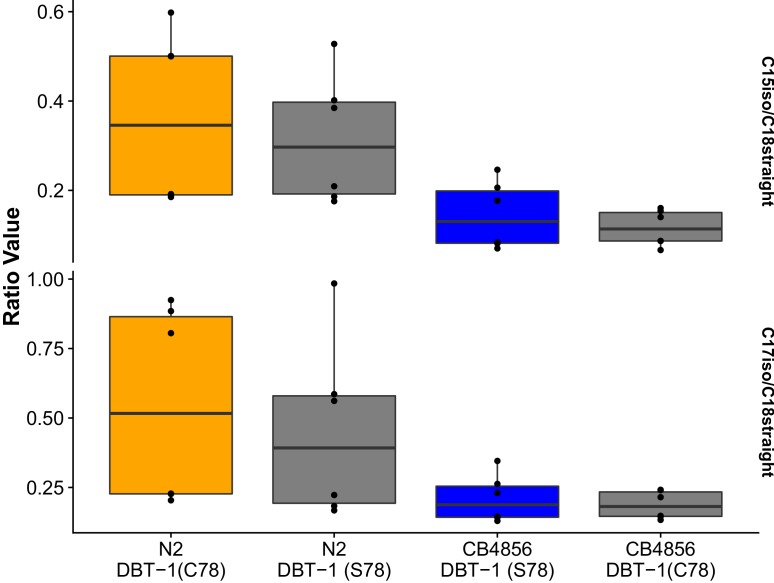
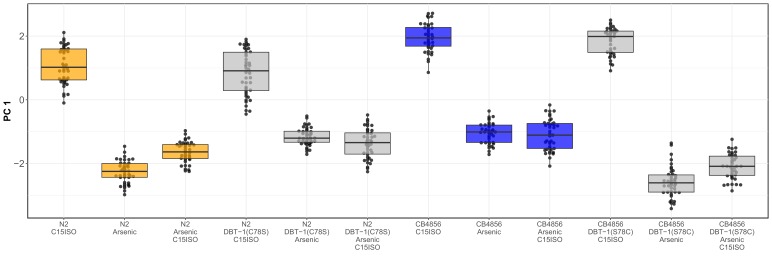
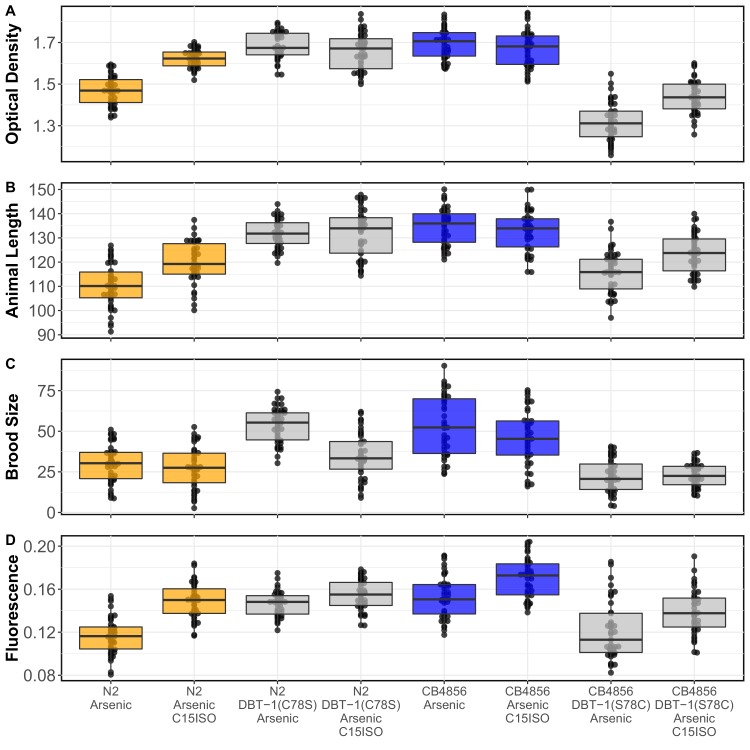
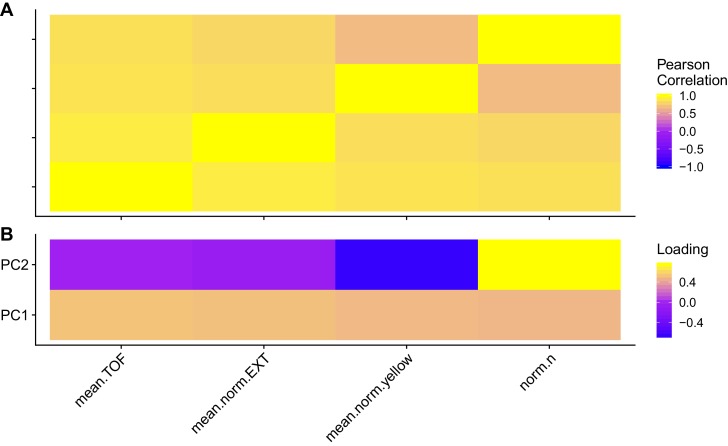
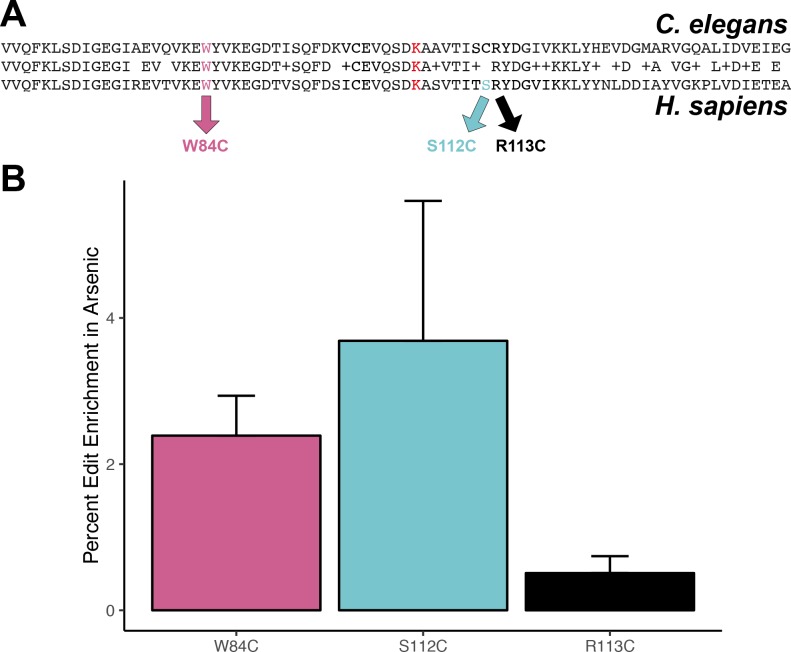
We find that variation in the gene underlies natural differences in responses to the toxin arsenic. This gene encodes the E2 subunit of the branched-chain α-keto acid dehydrogenase (BCKDH) complex, a core component of branched-chain amino acid (BCAA) metabolism. We causally linked a non-synonymous variant in the conserved lipoyl domain of DBT-1 to differential arsenic responses. Using targeted metabolomics and chemical supplementation, we demonstrate that differences in responses to arsenic are caused by variation in iso-branched chain fatty acids. Additionally, we show that levels of branched chain fatty acids in human cells are perturbed by arsenic treatment. This finding has broad implications for arsenic toxicity and for arsenic-focused chemotherapeutics across human populations. Our study implicates the BCKDH complex and BCAA metabolism in arsenic responses, demonstrating the power of natural genetic diversity to identify novel mechanisms by which environmental toxins affect organismal physiology.
This article has been through an editorial process in which the authors decide how to respond to the issues raised during peer review. The Reviewing Editor's assessment is that all the issues have been addressed (see decision letter).
我们发现基因的变异是砷毒素反应自然差异的基础。该基因编码支链α-酮酸脱氢酶(BCKDH)复合物的 E2 亚基,这是支链氨基酸(BCAA)代谢的核心组成部分。我们将 DBT-1 中保守的脂酰结构域中的非同义变体与砷反应的差异联系起来。通过靶向代谢组学和化学补充,我们证明了对砷的反应差异是由异支链脂肪酸的差异引起的。此外,我们还表明,砷处理会扰乱人类细胞中支链脂肪酸的水平。这一发现对砷毒性以及砷为重点的化疗药物在人类中的应用具有广泛的影响。我们的研究表明,BCKDH 复合物和 BCAA 代谢参与了砷反应,证明了天然遗传多样性在识别环境毒素影响生物体生理的新机制方面的力量。
本文经过编辑过程,作者决定如何处理同行评审期间提出的问题。审稿人的评估是所有问题都已得到解决(见评审意见)。