Department of Biochemistry and Molecular Biology (B) and Immunology, Faculty of Chemistry, University of Murcia, Campus of Espinardo, Regional Campus of International Excellence ''Campus Mare Nostrum'', P.O. Box 4021, 30100, Murcia, Spain.
Microb Cell Fact. 2019 Sep 4;18(1):151. doi: 10.1186/s12934-019-1202-1.
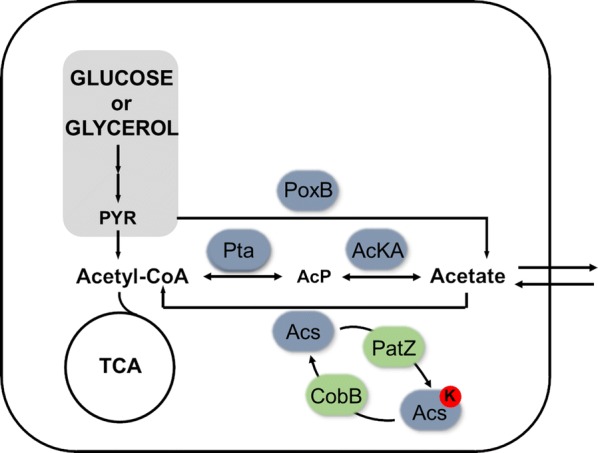
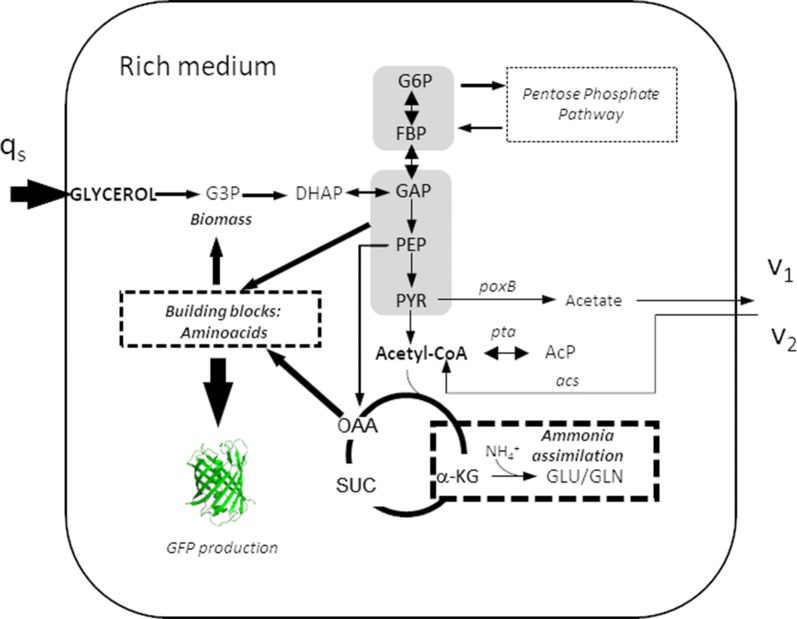
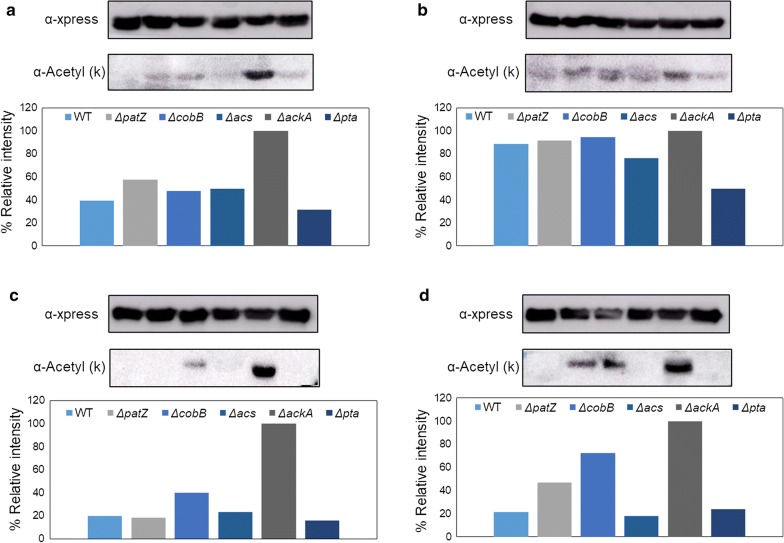
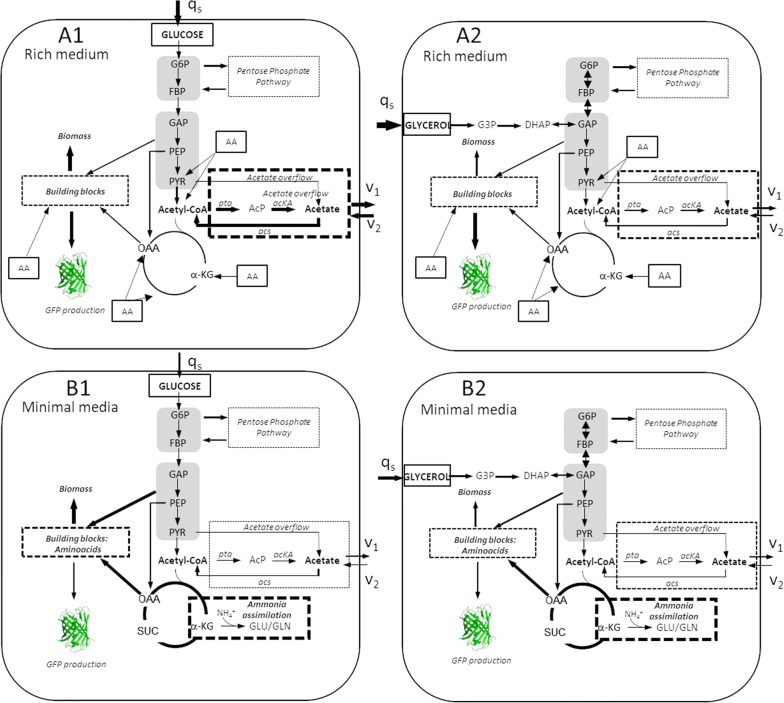
Escherichia coli (E. coli) is a bacteria that is widely employed in many industries for the production of high interest bio-products such as recombinant proteins. Nevertheless, the use of E. coli for recombinant protein production may entail some disadvantages such as acetate overflow. Acetate is accumulated under some culture conditions, involves a decrease in biomass and recombinant protein production, and its metabolism is related to protein lysine acetylation. Thereby, the carbon and nitrogen sources employed are relevant factors in cell host metabolism, and the study of the central metabolism of E. coli and its regulation is essential for optimizing the production of biomass and recombinant proteins. In this study, our aim was to find the most favourable conditions for carrying out recombinant protein production in E. coli BL21 using two different approaches, namely, manipulation of the culture media composition and the deletion of genes involved in acetate metabolism and Nε-lysine acetylation.
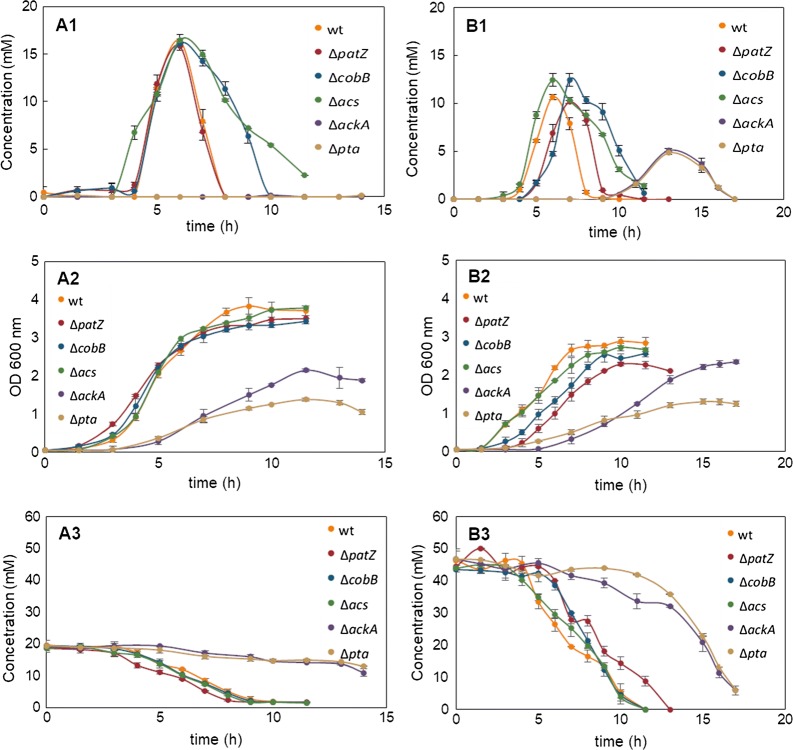
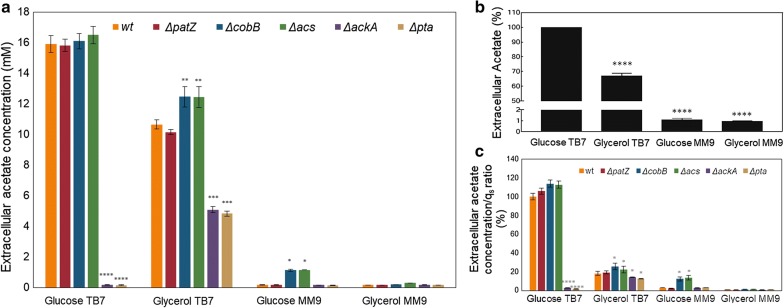
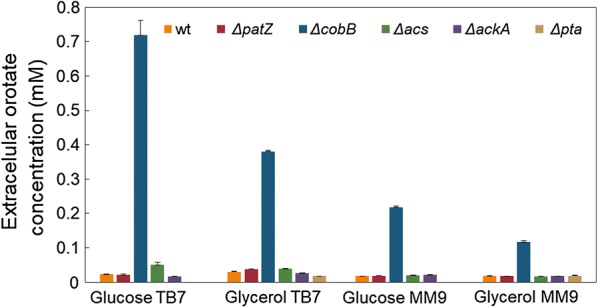
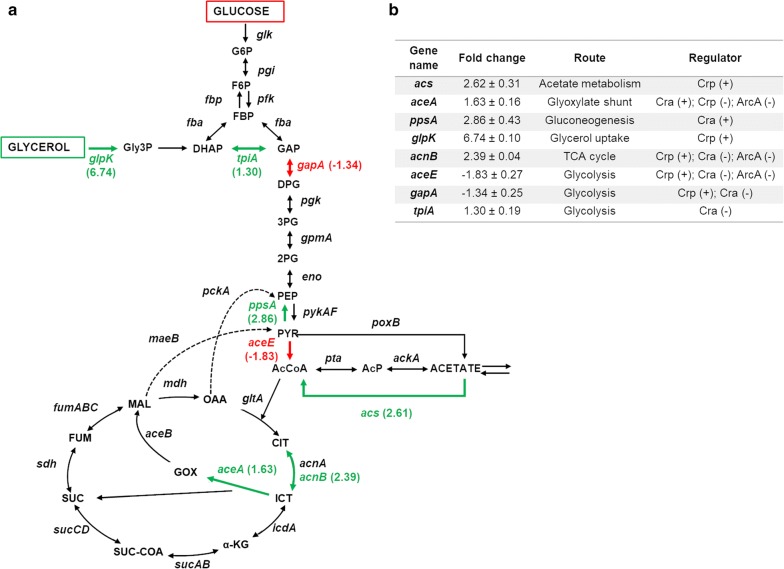
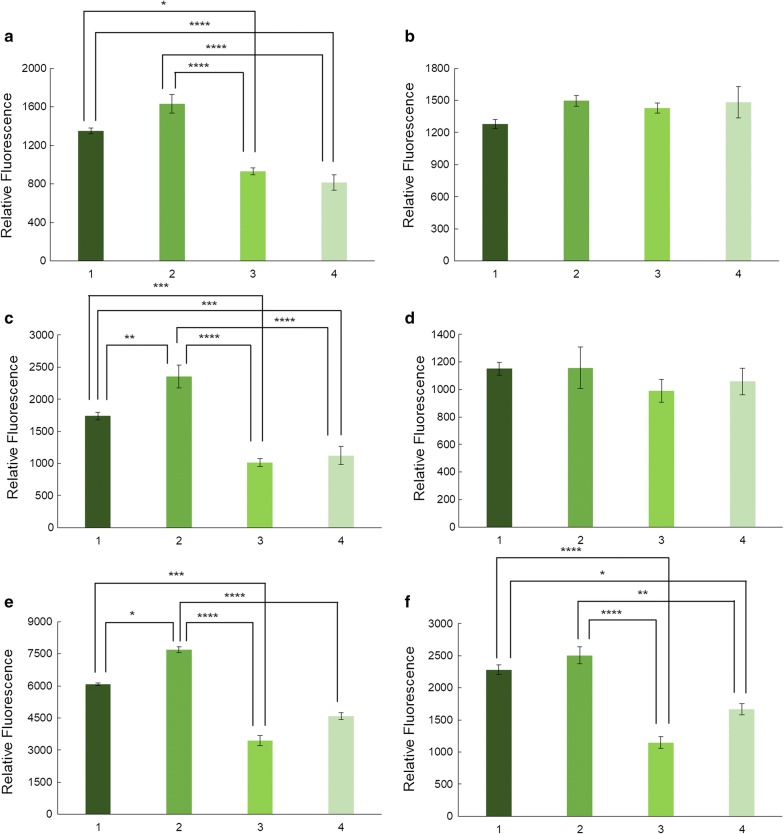
We evaluated protein overexpression in E. coli BL21 wt and five mutant strains involved in acetate metabolism (Δacs, ΔackA and Δpta) and lysine acetylation (ΔpatZ and ΔcobB) grown in minimal medium M9 (inorganic ammonium nitrogen source) and in complex TB7 medium (peptide-based nitrogen source) supplemented with glucose (PTS carbon source) or glycerol (non-PTS carbon source). We observed a dependence of recombinant protein production on acetate metabolism and the carbon and nitrogen source employed. The use of complex medium supplemented with glycerol as a carbon source entails an increase in protein production and an efficient use of resources, since is a sub-product of biodiesel synthesis. Furthermore, the deletion of the ackA gene results in a fivefold increase in protein production with respect to the wt strain and a reduction in acetate accumulation.
The results showed that the use of diverse carbon and nitrogen sources and acetate metabolism knockout strains can redirect E. coli carbon fluxes to different pathways and affect the final yield of the recombinant protein bioprocess. Thereby, we obtained a fivefold increase in protein production and an efficient use of the resources employing the most suitable strain and culture conditions.
大肠杆菌(E. coli)是一种在许多行业中广泛用于生产高价值生物制品(如重组蛋白)的细菌。然而,使用大肠杆菌生产重组蛋白可能存在一些缺点,例如乙酸盐溢出。在某些培养条件下,乙酸盐会积累,导致生物量和重组蛋白产量下降,其代谢与蛋白质赖氨酸乙酰化有关。因此,所使用的碳源和氮源是细胞宿主代谢的相关因素,研究大肠杆菌的中心代谢及其调控对于优化生物量和重组蛋白的生产至关重要。在这项研究中,我们的目的是通过两种不同的方法,即操纵培养基组成和删除与乙酸盐代谢和 Nε-赖氨酸乙酰化相关的基因,找到在大肠杆菌 BL21 中进行重组蛋白生产的最有利条件。
我们评估了在最小培养基 M9(无机铵氮源)和补充有葡萄糖(PTS 碳源)或甘油(非 PTS 碳源)的复杂 TB7 培养基(基于肽的氮源)中生长的大肠杆菌 BL21wt 和五个涉及乙酸盐代谢(Δacs、ΔackA 和 Δpta)和赖氨酸乙酰化(ΔpatZ 和 ΔcobB)的突变株中重组蛋白的过度表达。我们观察到重组蛋白的生产依赖于乙酸盐代谢和所使用的碳源和氮源。使用复杂培养基补充甘油作为碳源会增加蛋白产量并有效地利用资源,因为甘油是生物柴油合成的副产物。此外,敲除 ackA 基因可使蛋白产量相对于 wt 菌株增加五倍,并减少乙酸盐的积累。
结果表明,使用不同的碳源和氮源以及敲除乙酸盐代谢基因可以使大肠杆菌的碳通量重新定向到不同的途径,并影响重组蛋白生物过程的最终产量。因此,我们采用最合适的菌株和培养条件,使蛋白产量增加了五倍,并有效地利用了资源。