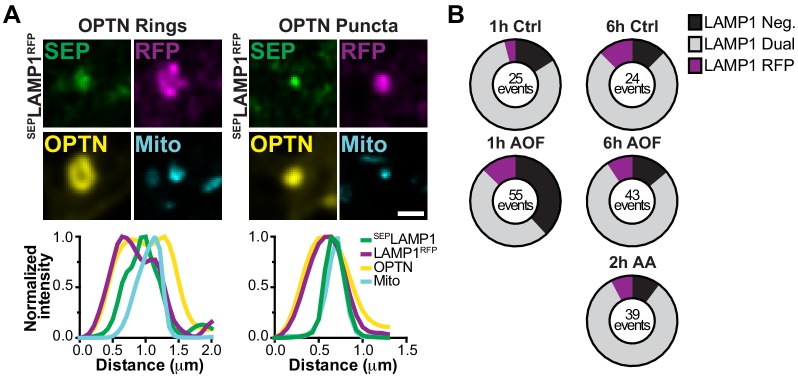
Evans Chantell S, Holzbaur Erika Lf
Department of Physiology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, United States.
Elife. 2020 Jan 14;9:e50260. doi: 10.7554/eLife.50260.
Mitophagy, the selective removal of damaged mitochondria, is thought to be critical to maintain neuronal homeostasis. Mutations of proteins in the pathway cause neurodegenerative diseases, suggesting defective mitochondrial turnover contributes to neurodegeneration. In primary rat hippocampal neurons, we developed a mitophagy induction paradigm where mild oxidative stress induced low levels of mitochondrial damage. Mitophagy-associated proteins were sequentially recruited to depolarized mitochondria followed by sequestration into autophagosomes. The localization of these mitophagy events had a robust somal bias. In basal and induced conditions, engulfed mitochondria remained in non-acidified organelles for hours to days, illustrating efficient autophagosome sequestration but delayed lysosomal fusion or acidification. Furthermore, expression of an ALS-linked mutation in the pathway disrupted mitochondrial network integrity and this effect was exacerbated by oxidative stress. Thus, age-related decline in neuronal health or expression of disease-associated mutations in the pathway may exacerbate the slow kinetics of neuronal mitophagy, leading to neurodegeneration.
线粒体自噬,即对受损线粒体的选择性清除,被认为对于维持神经元内环境稳定至关重要。该通路中蛋白质的突变会引发神经退行性疾病,这表明线粒体更新缺陷会导致神经退行性变。在原代大鼠海马神经元中,我们建立了一种线粒体自噬诱导模式,即轻度氧化应激诱导低水平的线粒体损伤。线粒体自噬相关蛋白会依次被招募到去极化的线粒体上,随后被隔离到自噬体中。这些线粒体自噬事件的定位具有明显的胞体偏向性。在基础状态和诱导状态下,被吞噬的线粒体在非酸化细胞器中会保留数小时至数天,这表明自噬体的隔离效率高,但溶酶体融合或酸化延迟。此外,该通路中与肌萎缩侧索硬化症(ALS)相关的突变表达会破坏线粒体网络的完整性,而氧化应激会加剧这种效应。因此,与年龄相关的神经元健康下降或该通路中疾病相关突变的表达可能会加剧神经元线粒体自噬的缓慢动力学,从而导致神经退行性变。