ESI, Biosciences, University of Exeter, Penryn, UK.
Department of Biochemistry, University of Otago, Dunedin, New Zealand.
Nature. 2020 Feb;578(7793):149-153. doi: 10.1038/s41586-020-1936-2. Epub 2020 Jan 22.
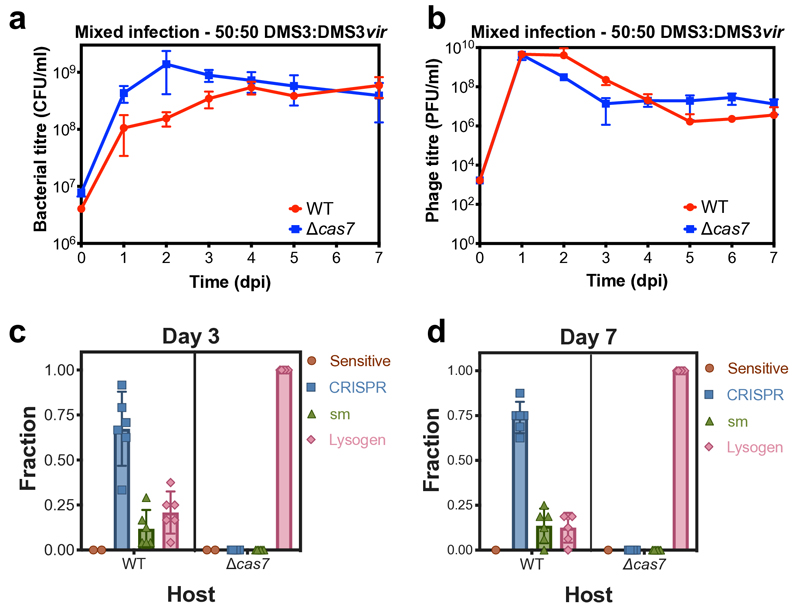
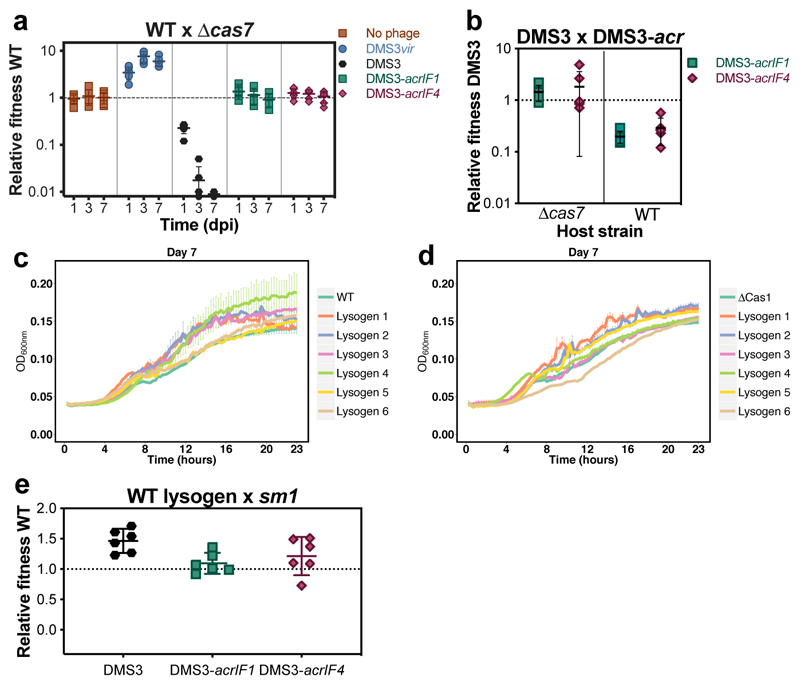
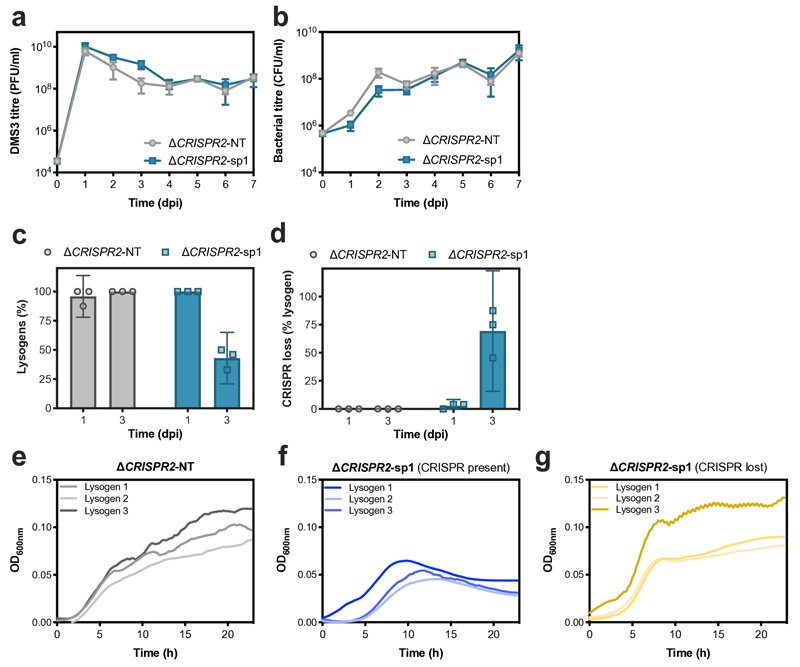
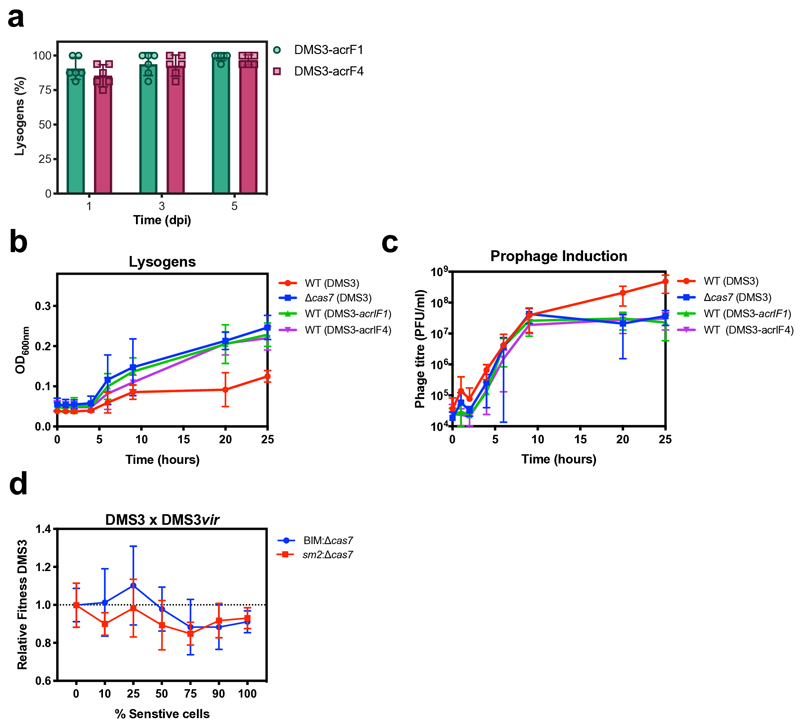
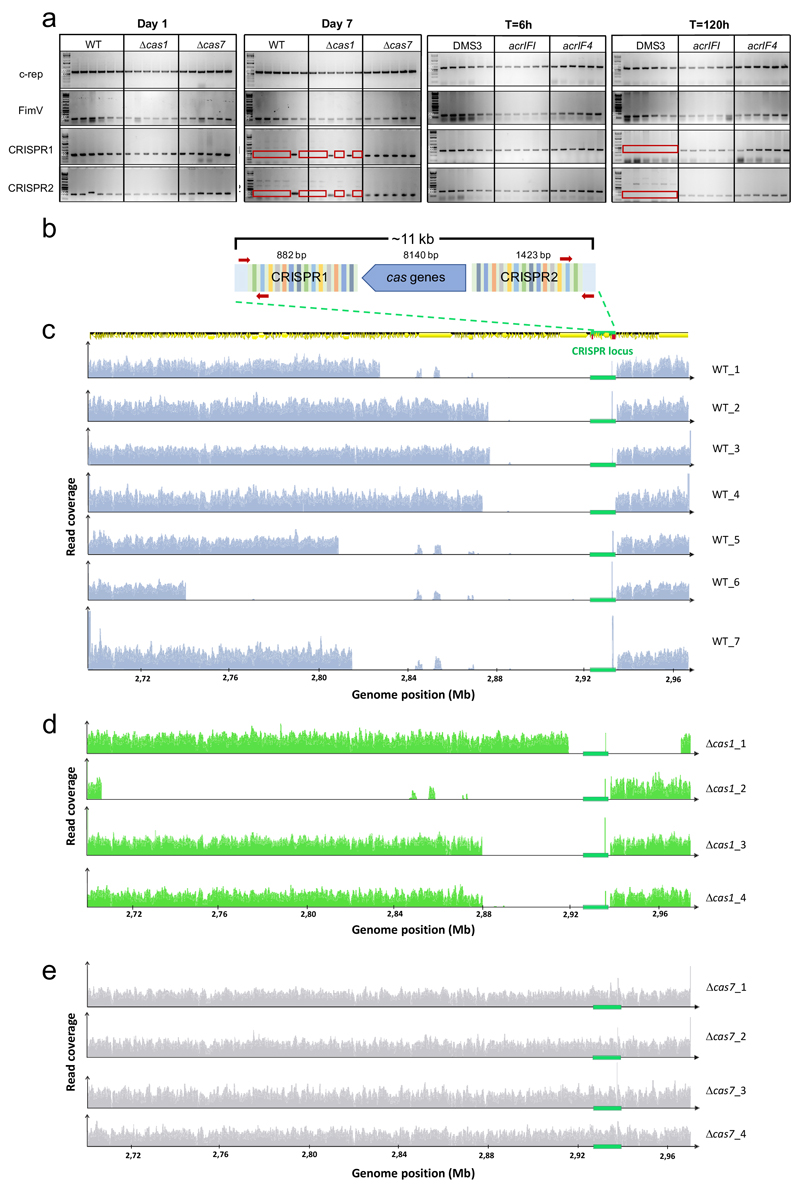
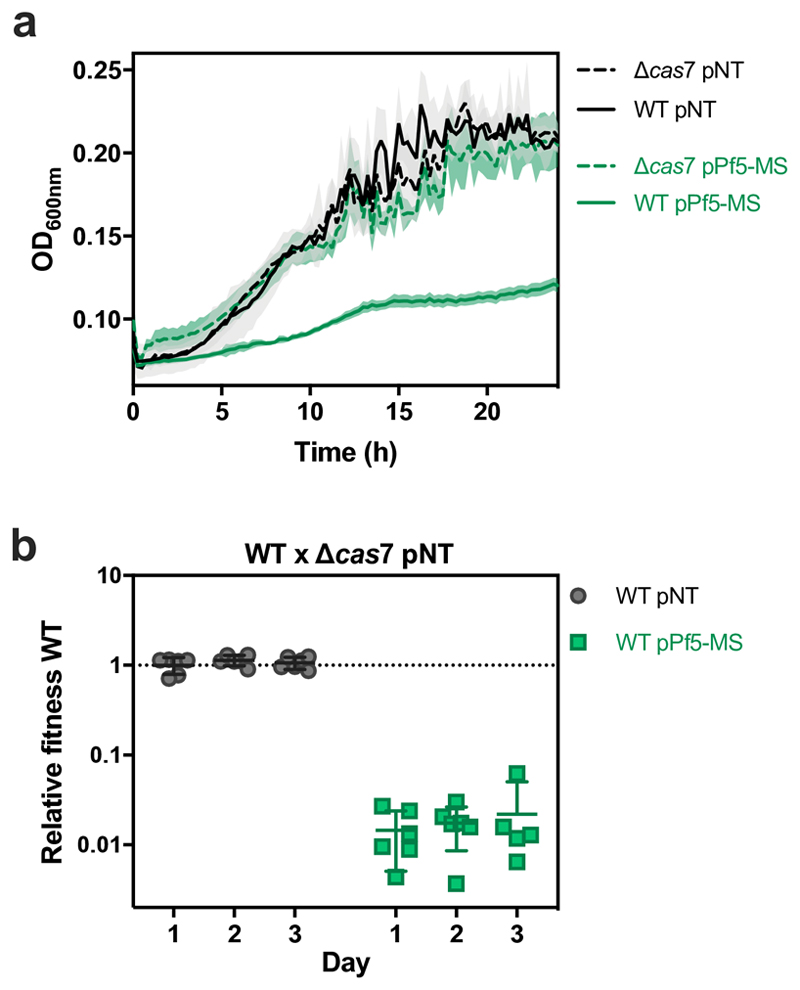
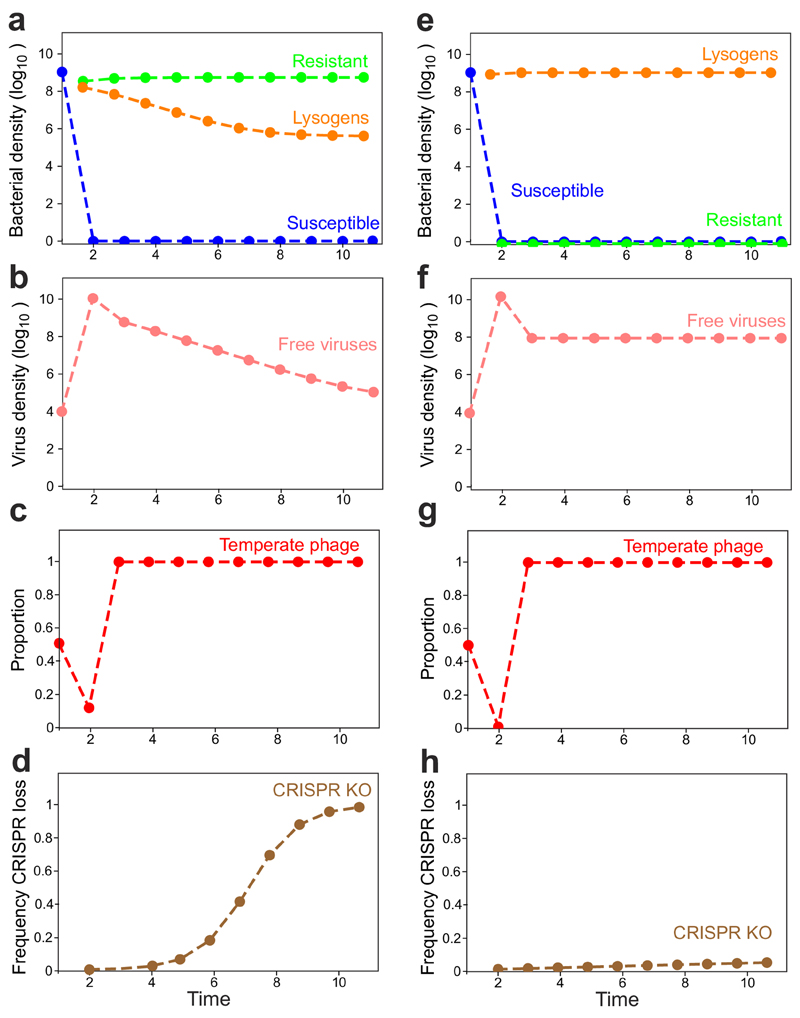
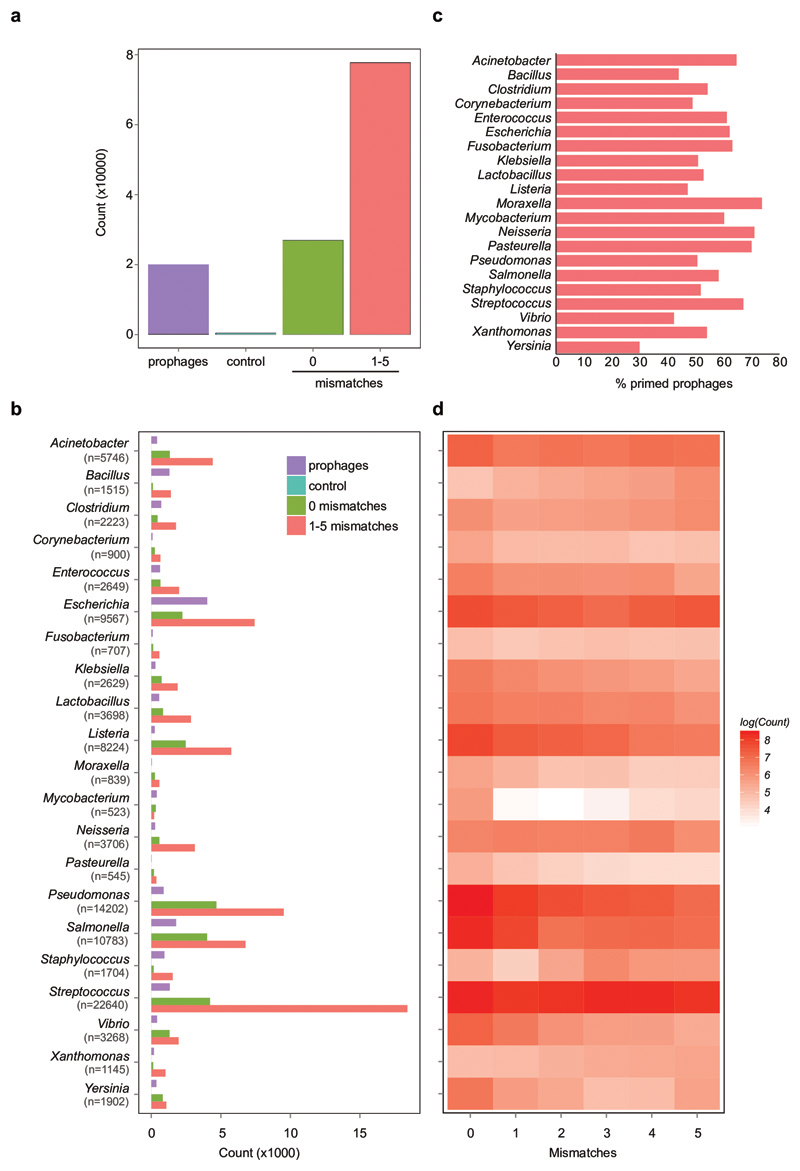
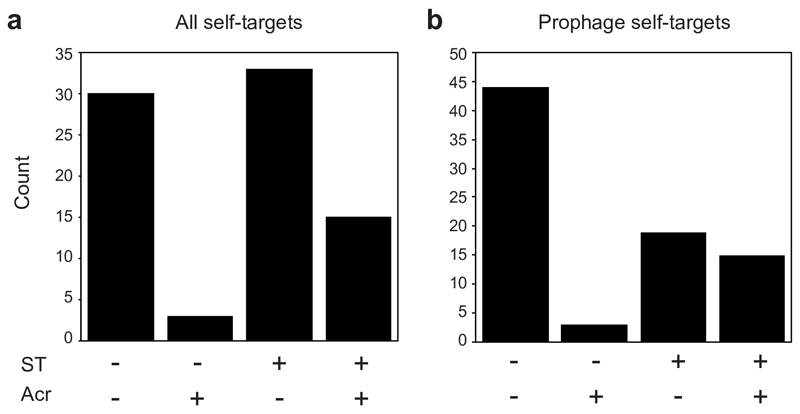
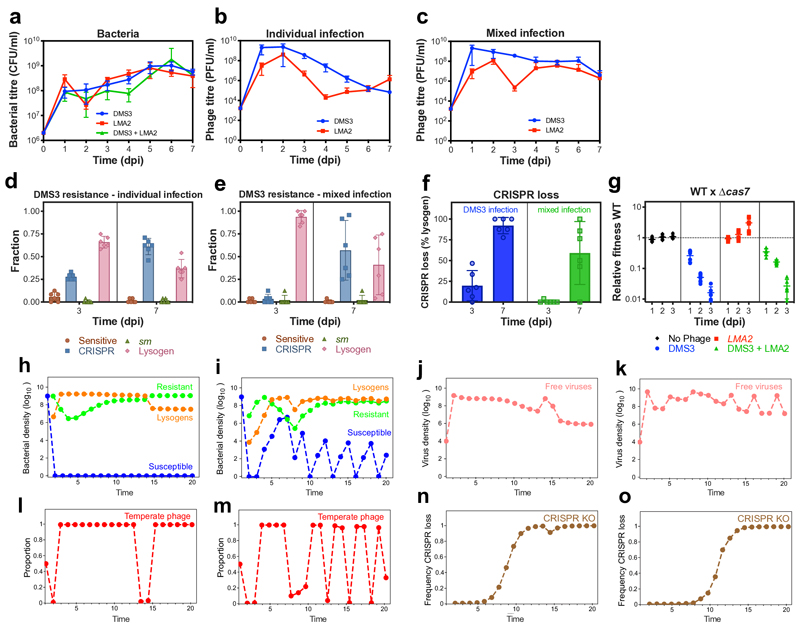
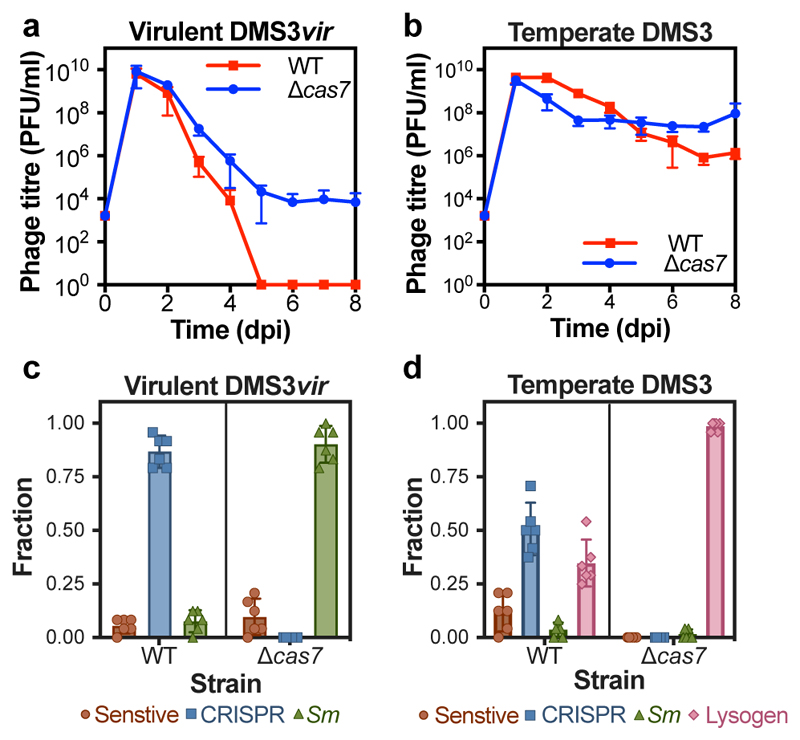
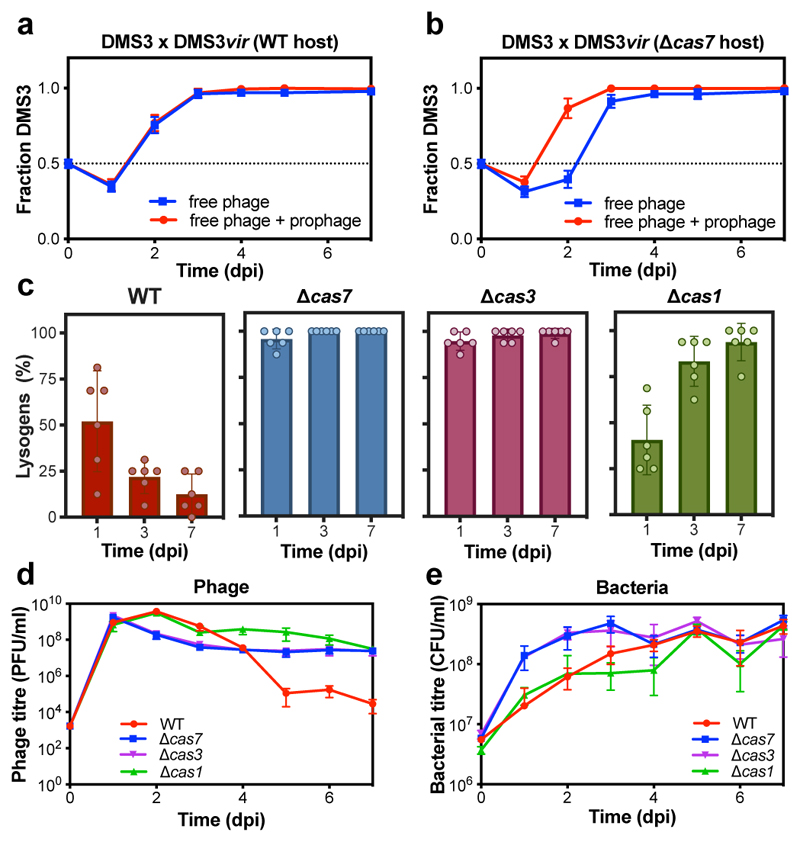
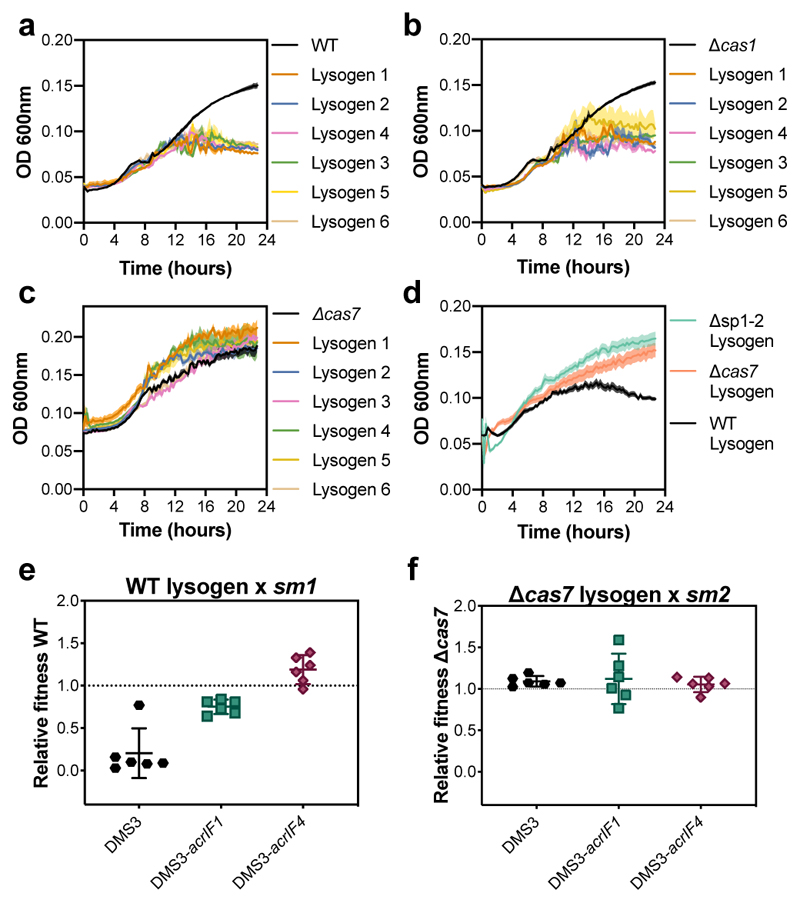
On infection of their host, temperate viruses that infect bacteria (bacteriophages; hereafter referred to as phages) enter either a lytic or a lysogenic cycle. The former results in lysis of bacterial cells and phage release (resulting in horizontal transmission), whereas lysogeny is characterized by the integration of the phage into the host genome, and dormancy (resulting in vertical transmission). Previous co-culture experiments using bacteria and mutants of temperate phages that are locked in the lytic cycle have shown that CRISPR-Cas systems can efficiently eliminate the invading phages. Here we show that, when challenged with wild-type temperate phages (which can become lysogenic), type I CRISPR-Cas immune systems cannot eliminate the phages from the bacterial population. Furthermore, our data suggest that, in this context, CRISPR-Cas immune systems are maladaptive to the host, owing to the severe immunopathological effects that are brought about by imperfect matching of spacers to the integrated phage sequences (prophages). These fitness costs drive the loss of CRISPR-Cas from bacterial populations, unless the phage carries anti-CRISPR (acr) genes that suppress the immune system of the host. Using bioinformatics, we show that this imperfect targeting is likely to occur frequently in nature. These findings help to explain the patchy distribution of CRISPR-Cas immune systems within and between bacterial species, and highlight the strong selective benefits of phage-encoded acr genes for both the phage and the host under these circumstances.
当感染其宿主时,感染细菌的温和噬菌体(以下简称噬菌体)进入裂解或溶原周期。前者导致细菌细胞裂解和噬菌体释放(导致水平传播),而溶原性的特征是噬菌体整合到宿主基因组中,并处于休眠状态(导致垂直传播)。以前使用细菌和处于裂解周期的温和噬菌体突变体的共培养实验表明,CRISPR-Cas 系统可以有效地消除入侵的噬菌体。在这里,我们表明,当受到野生型温和噬菌体(可能成为溶原性噬菌体)的挑战时,I 型 CRISPR-Cas 免疫系统无法从细菌群体中消除噬菌体。此外,我们的数据表明,在这种情况下,CRISPR-Cas 免疫系统对宿主是适应不良的,这是由于间隔区与整合的噬菌体序列(原噬菌体)不完全匹配所带来的严重免疫病理效应。这些适应不良的代价导致 CRISPR-Cas 从细菌群体中丢失,除非噬菌体携带抗 CRISPR(acr)基因,这些基因可以抑制宿主的免疫系统。我们利用生物信息学方法表明,这种不完全靶向在自然界中很可能经常发生。这些发现有助于解释 CRISPR-Cas 免疫系统在细菌种内和种间的分布不均,并强调了在这种情况下,噬菌体编码的 acr 基因对噬菌体和宿主都具有很强的选择优势。