Biochemie-Zentrum der Universität Heidelberg, Heidelberg, Germany.
Division of Redox Regulation, DKFZ-ZMBH Alliance, German Cancer Research Center (DKFZ), Heidelberg, Germany.
Elife. 2020 Jan 31;9:e53227. doi: 10.7554/eLife.53227.
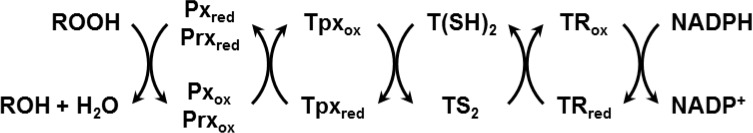
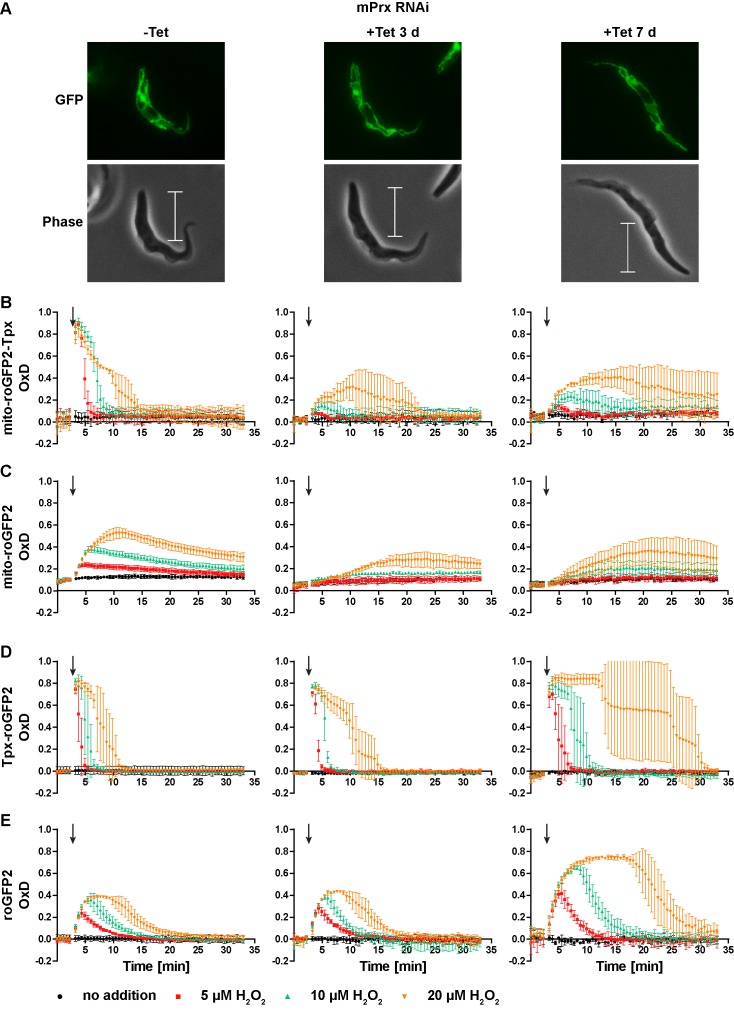
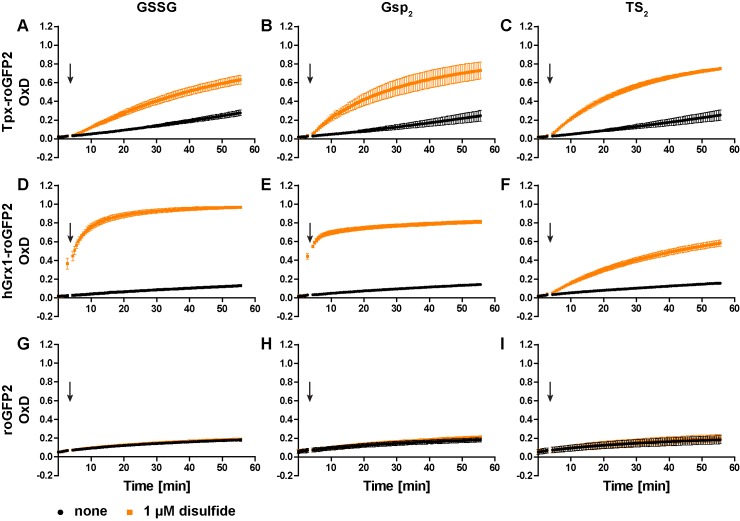
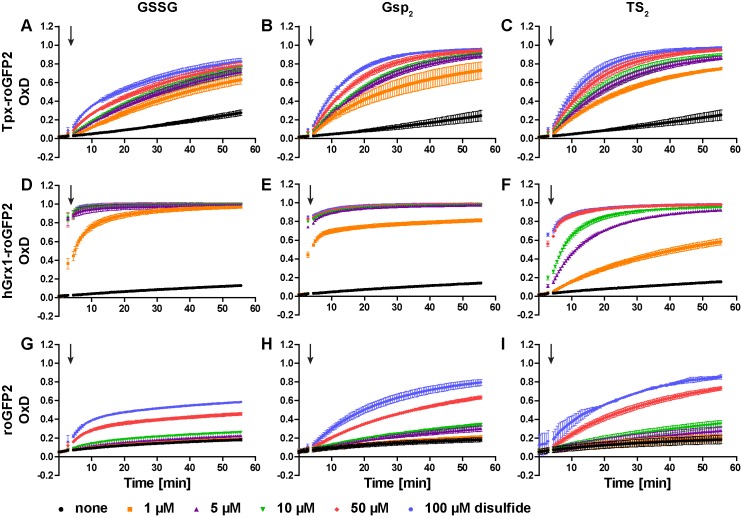
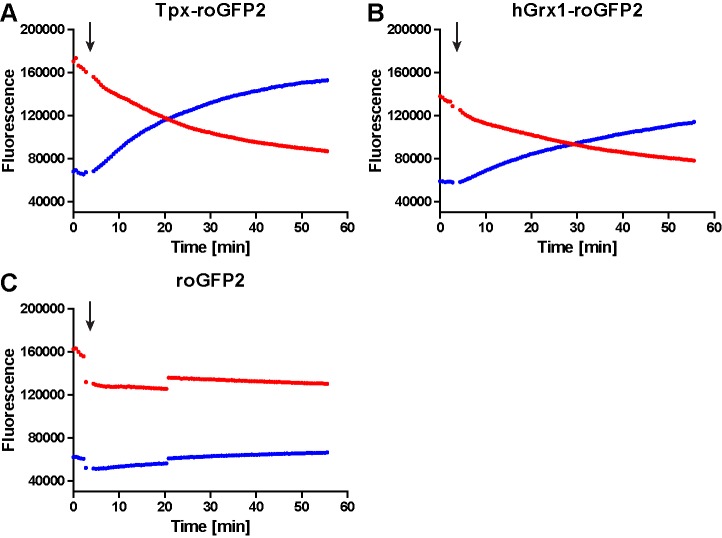
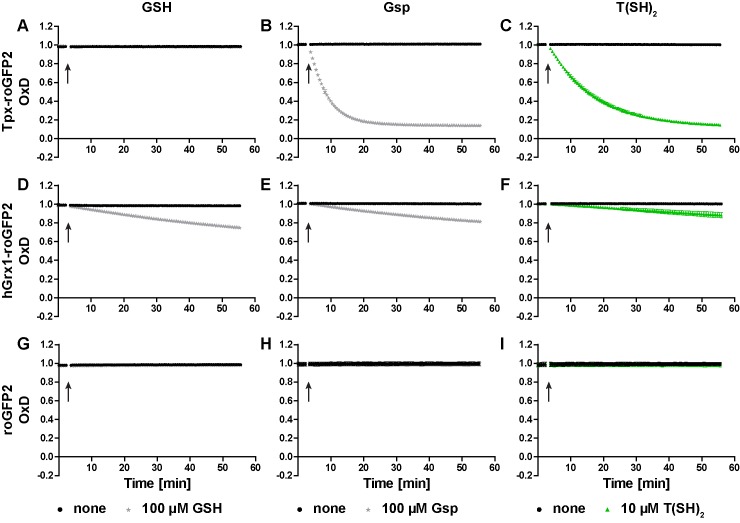
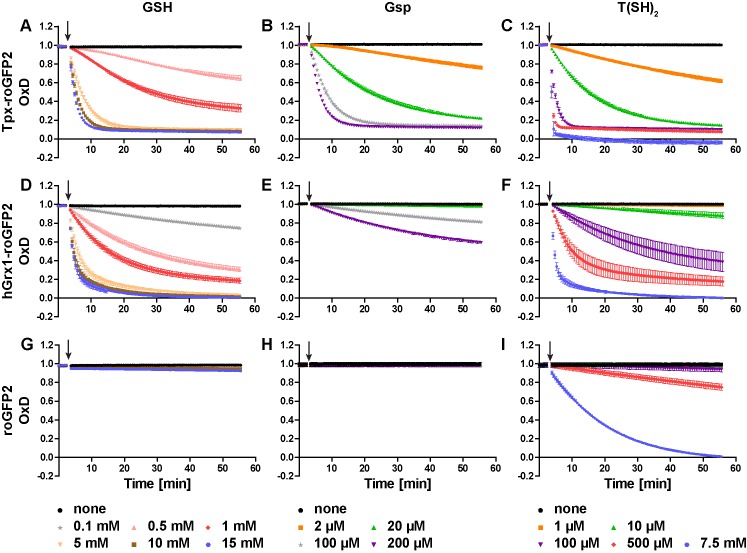
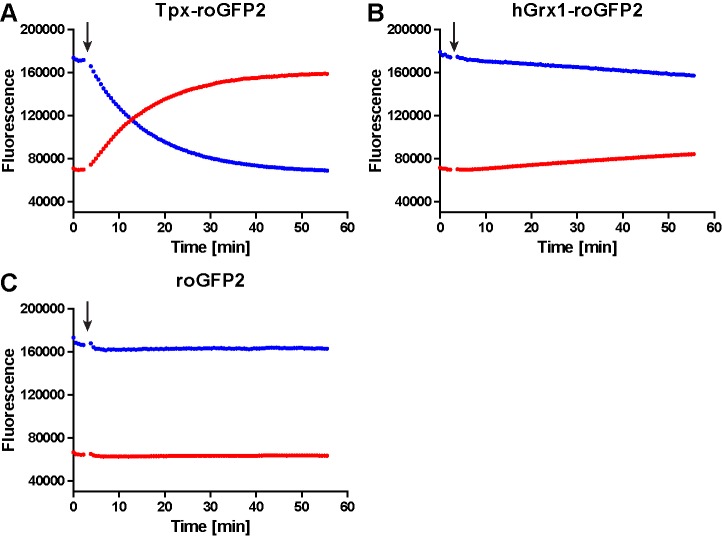
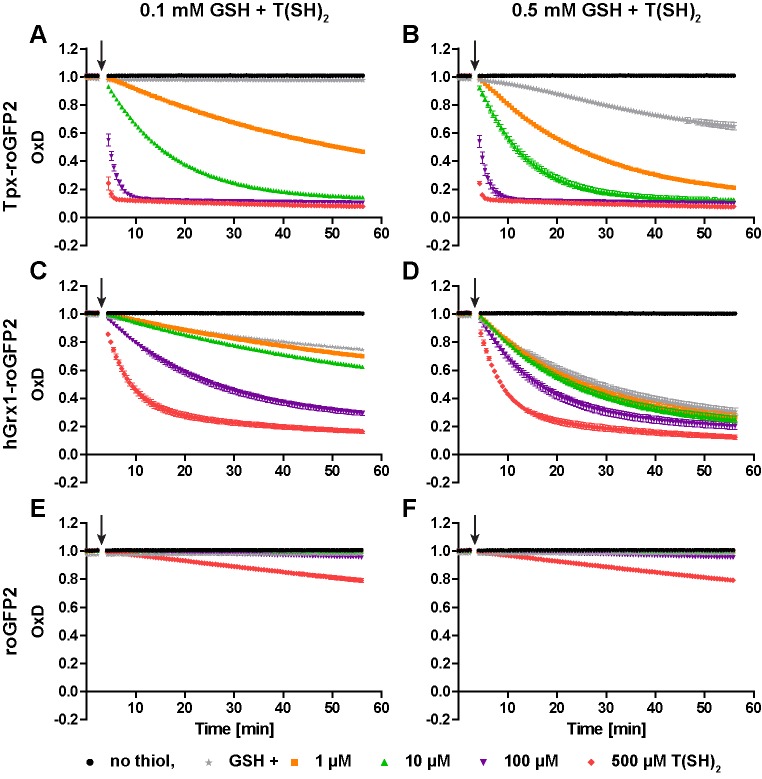
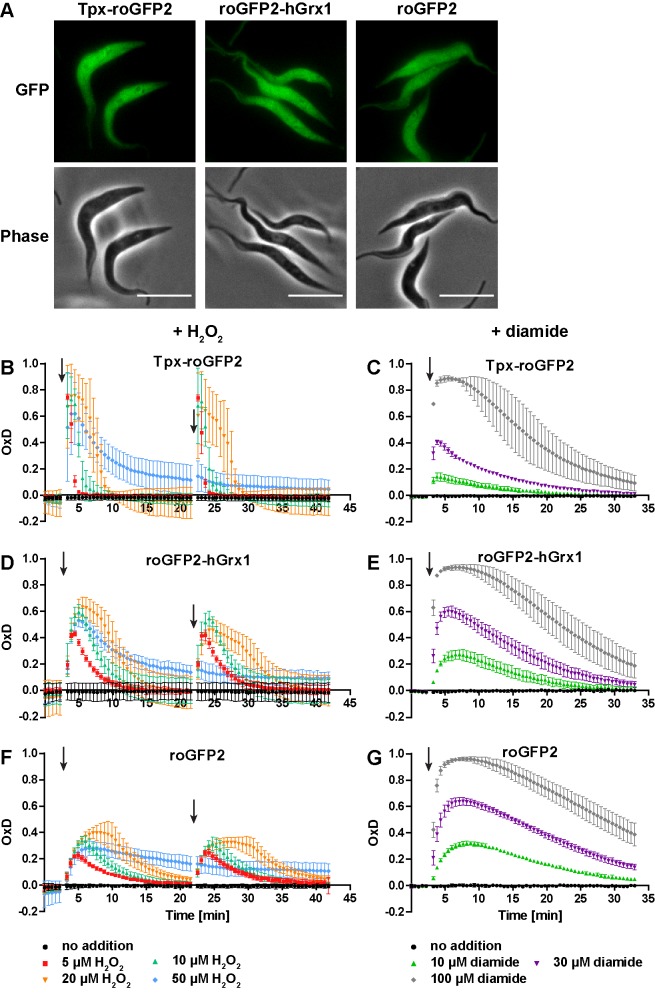
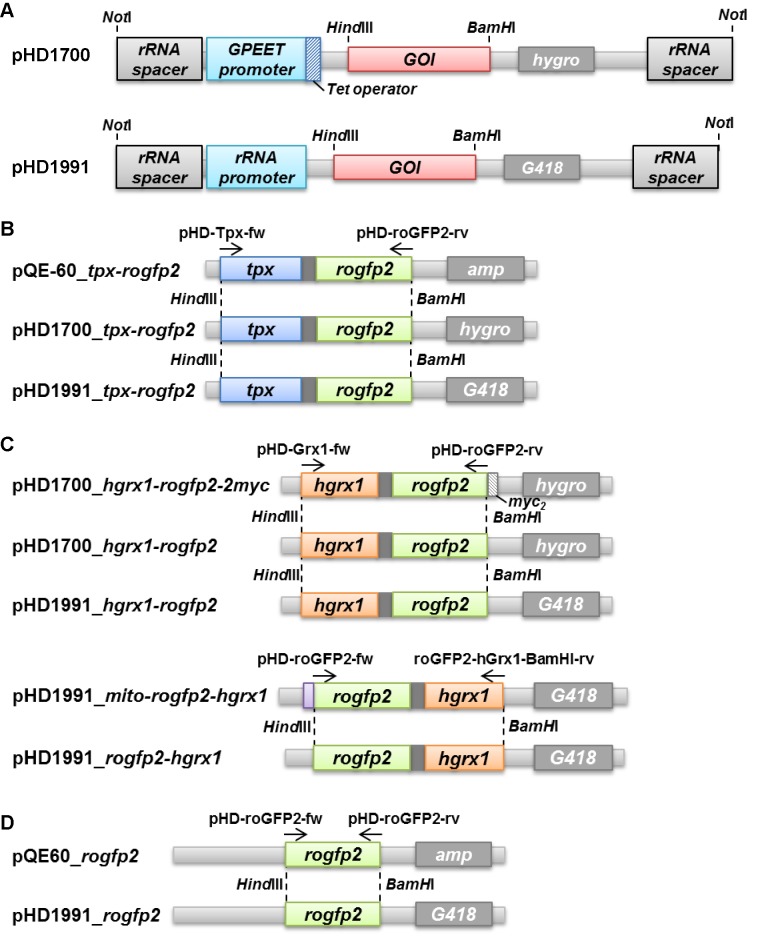
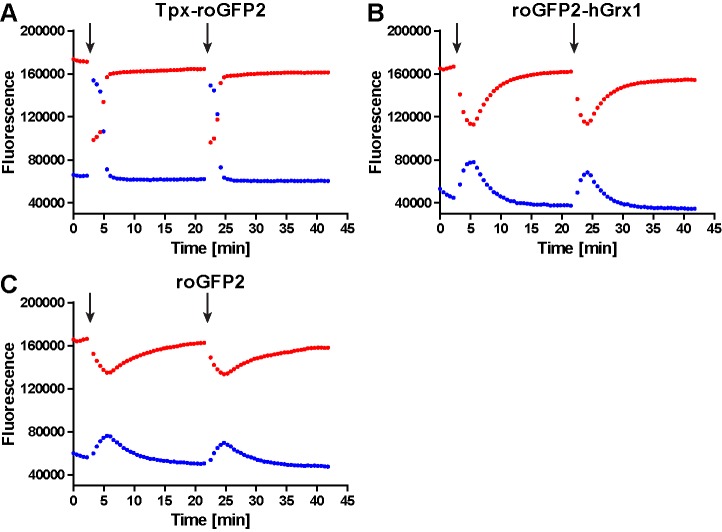
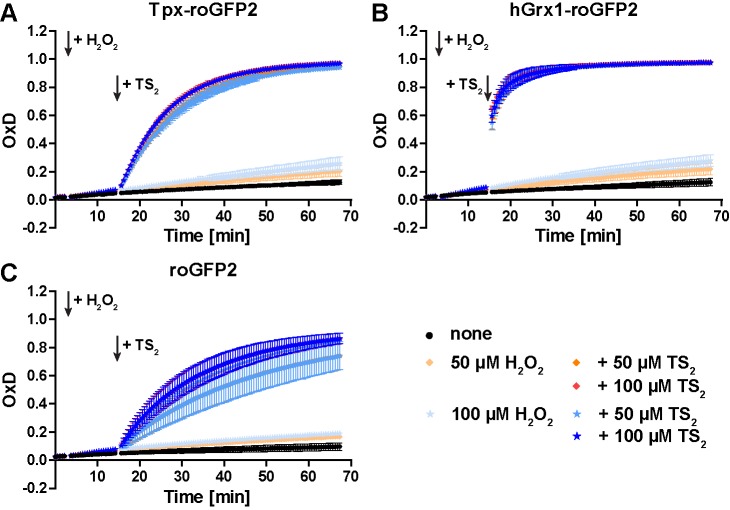
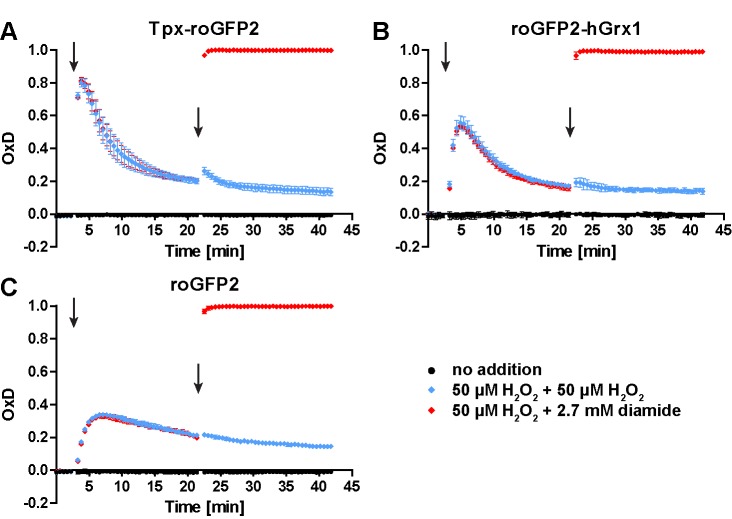
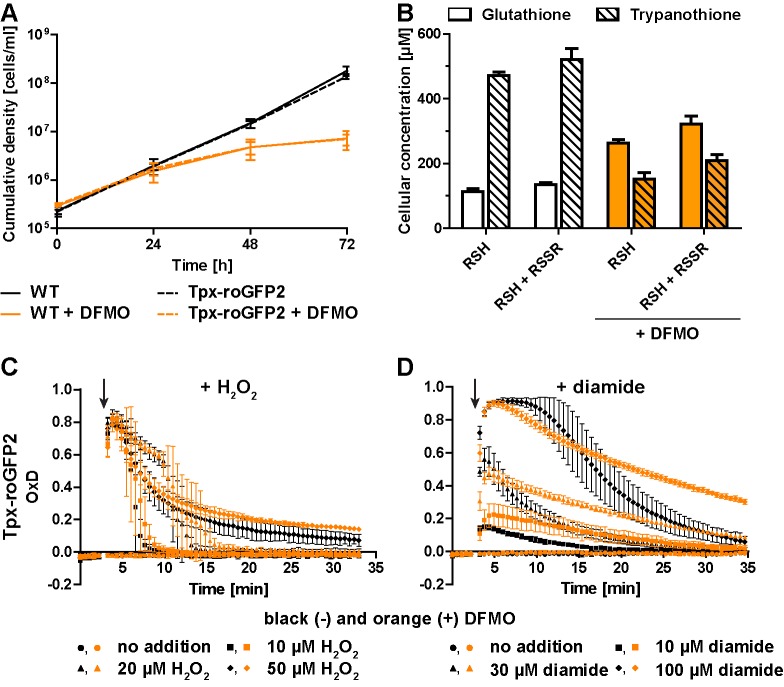
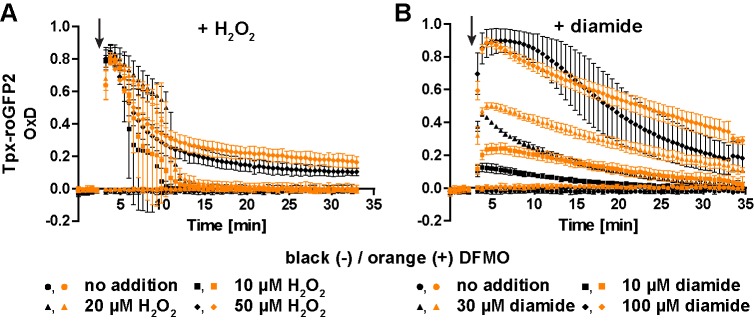
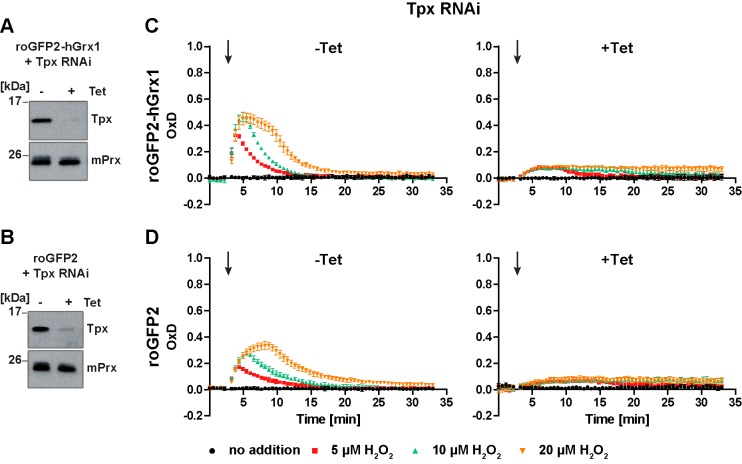
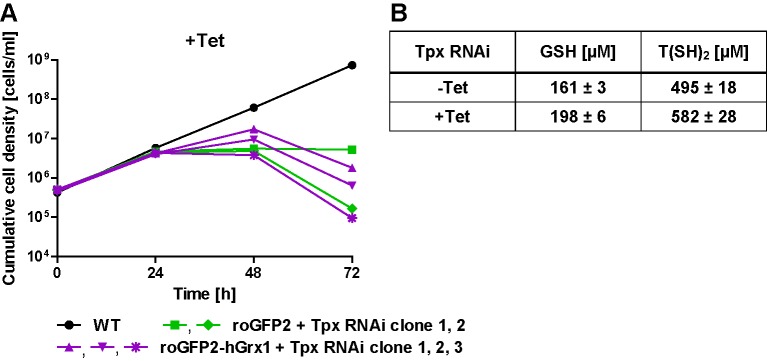
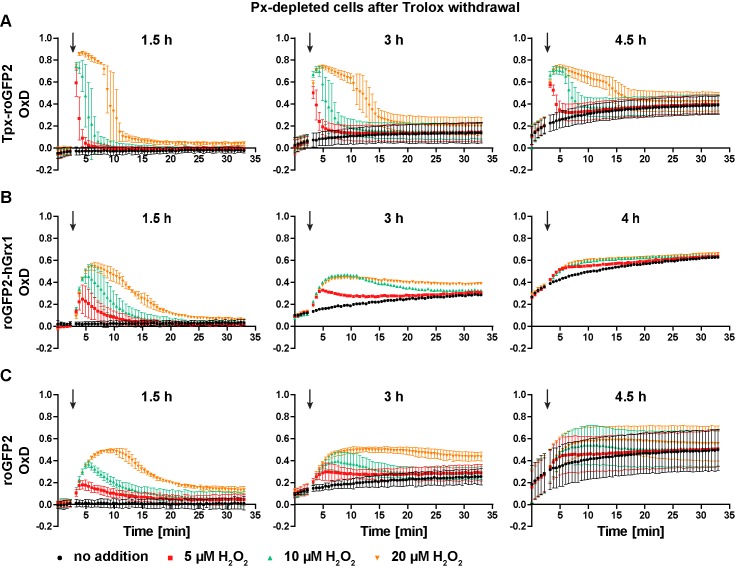
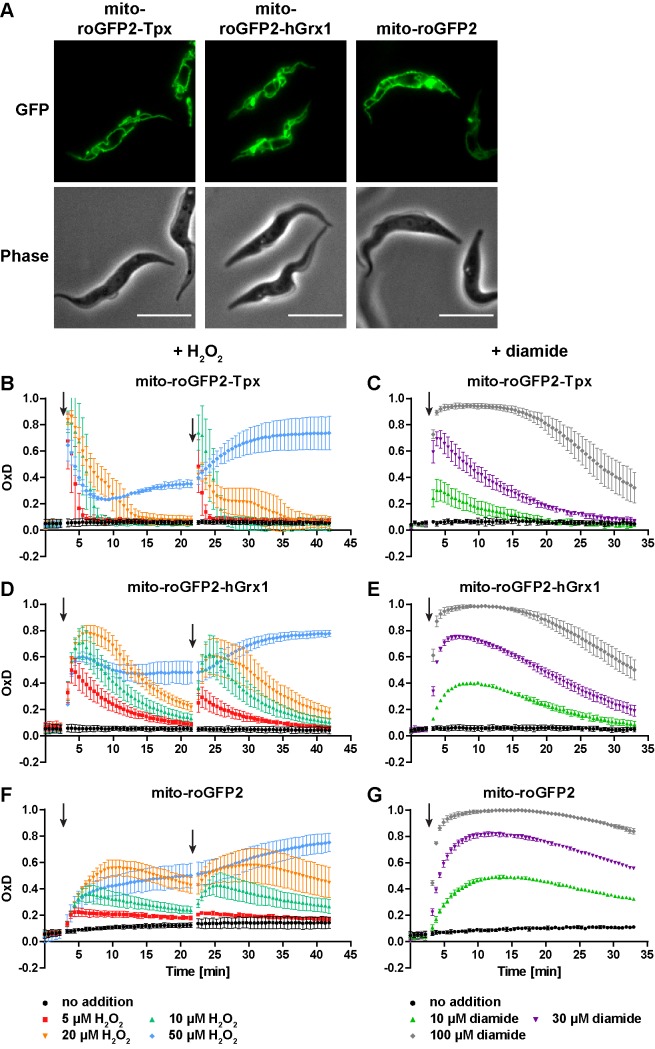
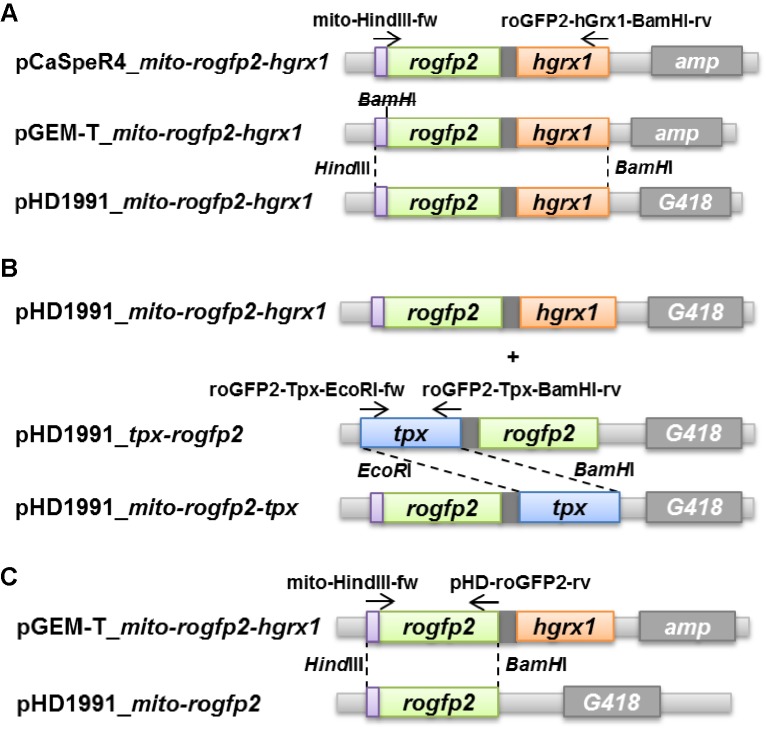
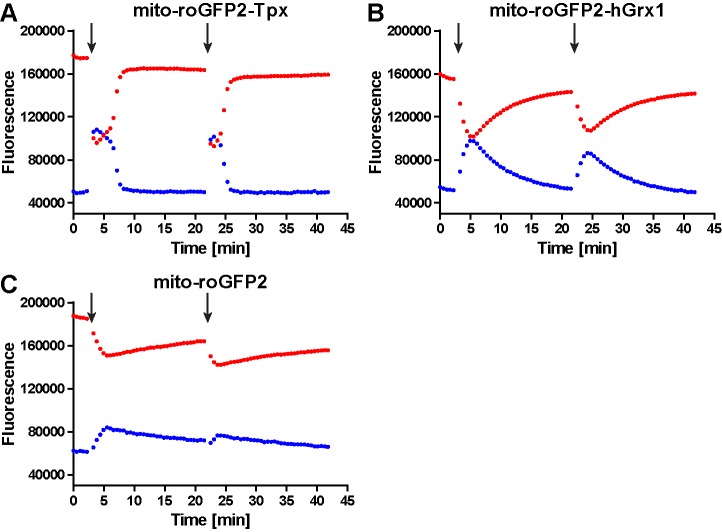
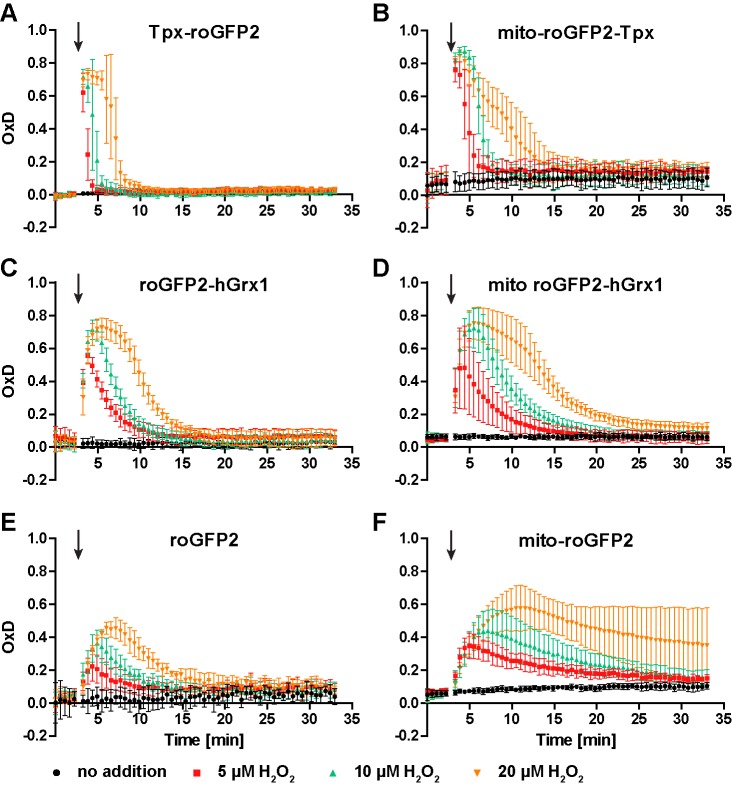
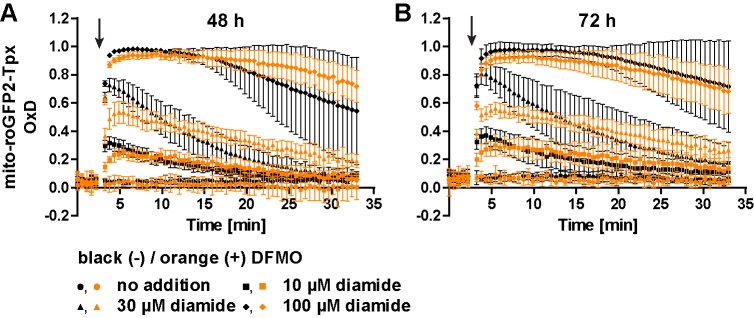
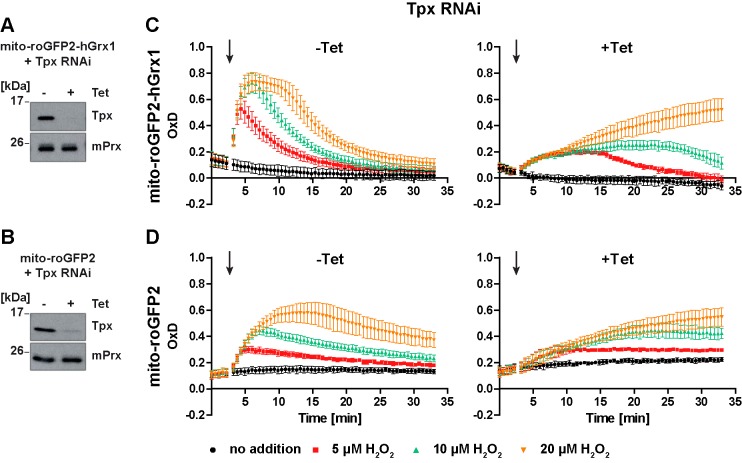
Trypanosomes have a trypanothione redox metabolism that provides the reducing equivalents for numerous essential processes, most being mediated by tryparedoxin (Tpx). While the biosynthesis and reduction of trypanothione are cytosolic, the molecular basis of the thiol redox homeostasis in the single mitochondrion of these parasites has remained largely unknown. Here we expressed Tpx-roGFP2, roGFP2-hGrx1 or roGFP2 in either the cytosol or mitochondrion of . We show that the novel Tpx-roGFP2 is a superior probe for the trypanothione redox couple and that the mitochondrial matrix harbors a trypanothione system. Inhibition of trypanothione biosynthesis by the anti-trypanosomal drug Eflornithine impairs the ability of the cytosol and mitochondrion to cope with exogenous oxidative stresses, indicating a direct link between both thiol systems. Tpx depletion abolishes the cytosolic, but only partially affects the mitochondrial sensor response to HO. This strongly suggests that the mitochondrion harbors some Tpx and, another, as yet unidentified, oxidoreductase.
锥虫具有一种转酮醇酶还原代谢,为许多重要过程提供还原当量,其中大多数过程由硫氧还蛋白(Tpx)介导。虽然转酮醇酶的生物合成和还原是在细胞质中进行的,但这些寄生虫的单个线粒体中硫醇氧化还原动态平衡的分子基础在很大程度上仍是未知的。在这里,我们在锥虫的细胞质或线粒体中表达了 Tpx-roGFP2、roGFP2-hGrx1 或 roGFP2。我们表明,新型 Tpx-roGFP2 是转酮醇酶氧化还原偶联的优越探针,并且线粒体基质中存在转酮醇酶系统。抗锥虫药物 Eflornithine 抑制转酮醇酶的生物合成会损害细胞质和线粒体应对外源性氧化应激的能力,表明这两个硫醇系统之间存在直接联系。Tpx 耗竭会破坏细胞质中的传感器反应,但仅部分影响线粒体对 HO 的传感器反应。这强烈表明线粒体中存在一些 Tpx 和另一种尚未确定的氧化还原酶。