Center for Precision Genome Editing and Genetic Technologies for Biomedicine, Institute of Gene Biology, Russian Academy of Sciences, 34/5 Vavilov St., Moscow, 119334, Russia.
Department of the Control of Genetic Processes, Institute of Gene Biology, Russian Academy of Sciences, 34/5 Vavilov St., Moscow, 119334, Russia.
Sci Rep. 2020 Feb 14;10(1):2677. doi: 10.1038/s41598-020-59459-5.
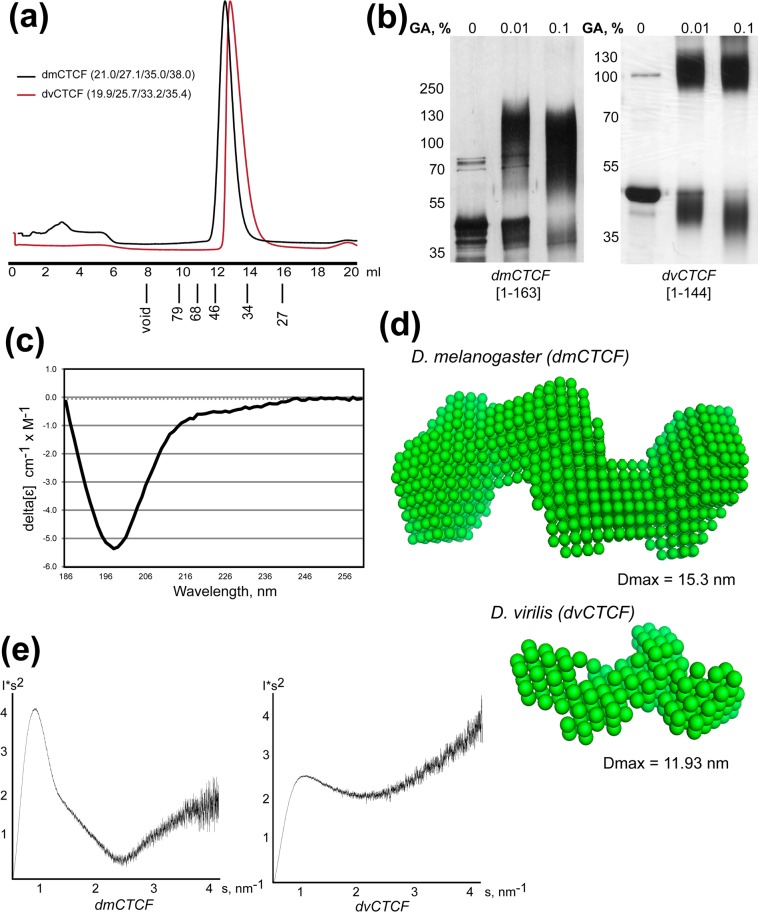
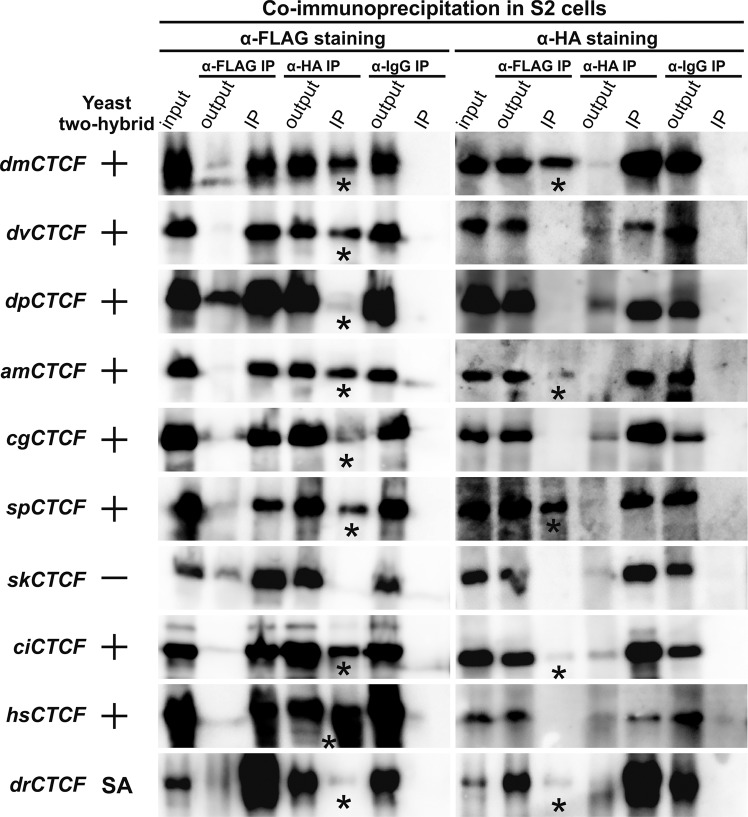
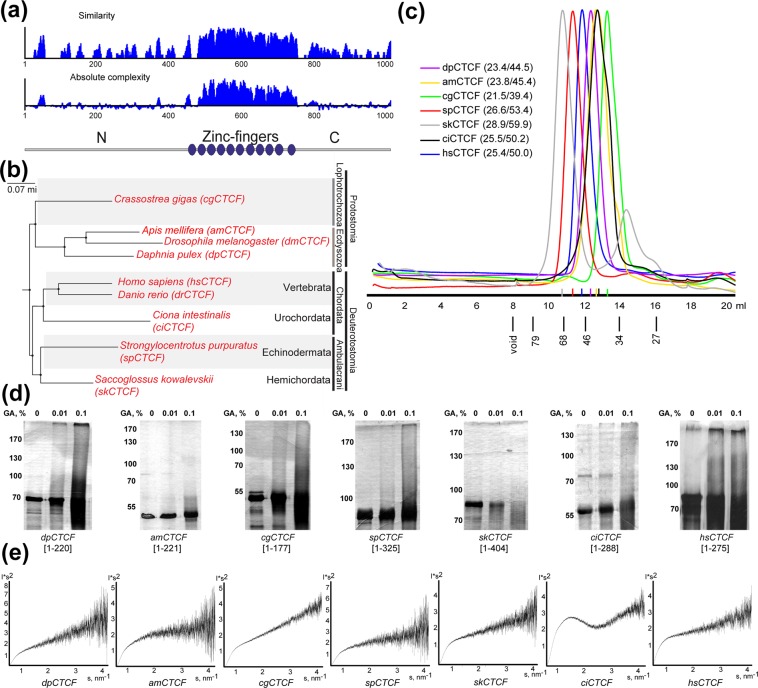
CTCF is the main architectural protein found in most of the examined bilaterian organisms. The cluster of the C2H2 zinc-finger domains involved in recognition of long DNA-binding motif is only part of the protein that is evolutionarily conserved, while the N-terminal domain (NTD) has different sequences. Here, we performed biophysical characterization of CTCF NTDs from various species representing all major phylogenetic clades of higher metazoans. With the exception of Drosophilides, the N-terminal domains of CTCFs show an unstructured organization and absence of folded regions in vitro. In contrast, NTDs of Drosophila melanogaster and virilis CTCFs contain unstructured folded regions that form tetramers and dimers correspondingly in vitro. Unexpectedly, most NTDs are able to self-associate in the yeast two-hybrid and co-immunoprecipitation assays. These results suggest that NTDs of CTCFs might contribute to the organization of CTCF-mediated long-distance interactions and chromosomal architecture.
CTCF 是在大多数被研究的两侧对称生物中发现的主要结构蛋白。参与识别长 DNA 结合基序的 C2H2 锌指结构域簇只是进化上保守的蛋白质的一部分,而 N 端结构域(NTD)具有不同的序列。在这里,我们对来自不同物种的 CTCF NTDs 进行了生物物理特性分析,这些物种代表了高等后生动物的所有主要系统发育进化枝。除了 Drosophilides 之外,CTCF 的 N 端结构域在体外表现出无规卷曲的组织和无折叠区域。相比之下,黑腹果蝇和 virilis CTCF 的 NTD 含有无规卷曲的折叠区域,在体外相应地形成四聚体和二聚体。出乎意料的是,大多数 NTDs 能够在酵母双杂交和共免疫沉淀实验中自我缔合。这些结果表明,CTCF 的 NTD 可能有助于 CTCF 介导的长距离相互作用和染色体结构的组织。