Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02142
Genetics. 2020 Mar;214(3):543-560. doi: 10.1534/genetics.119.301292.
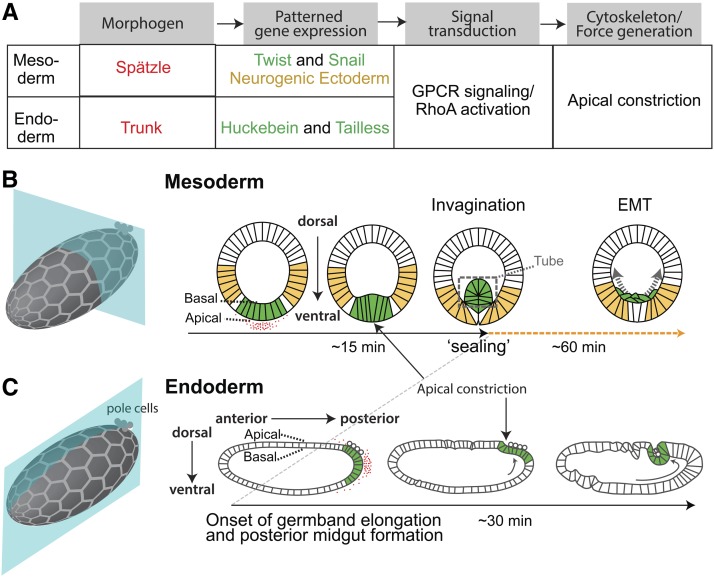
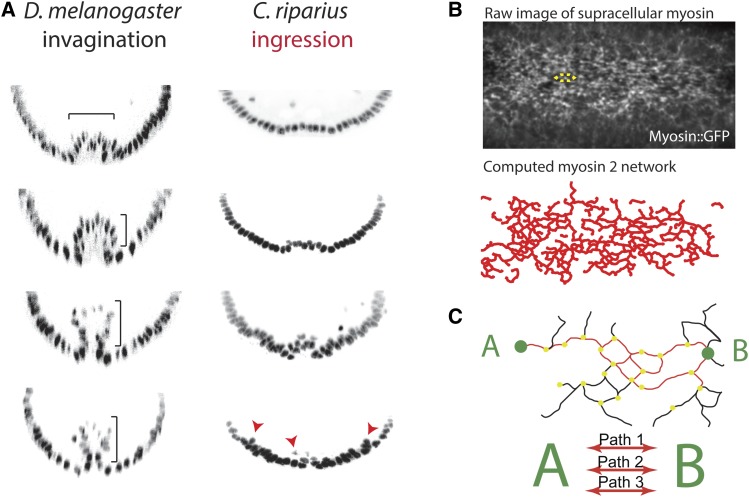
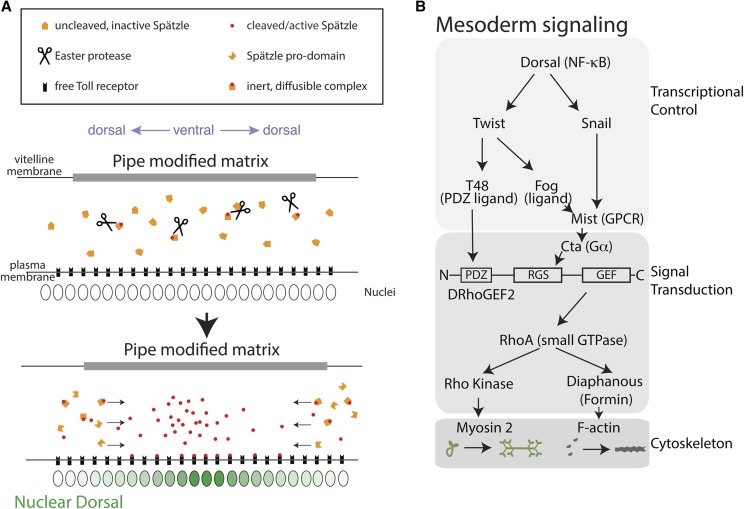
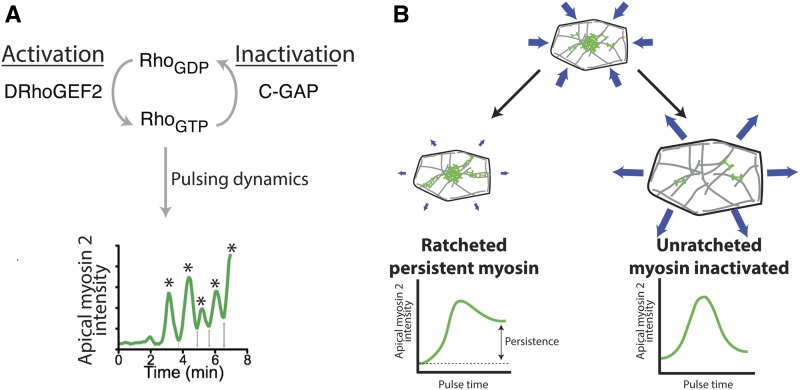
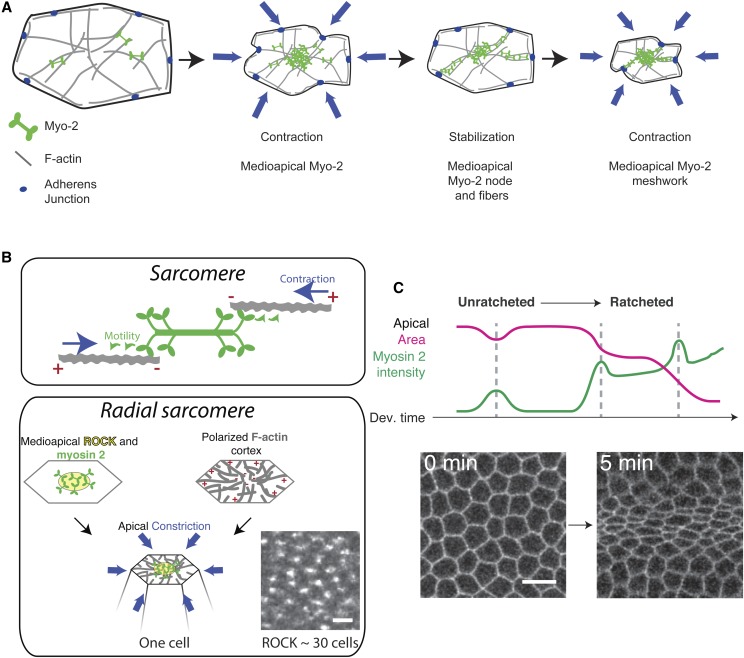
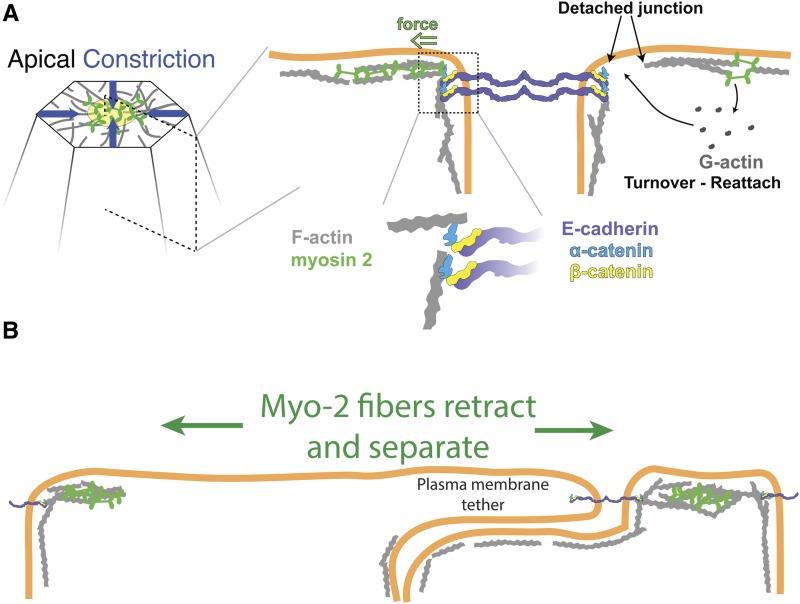
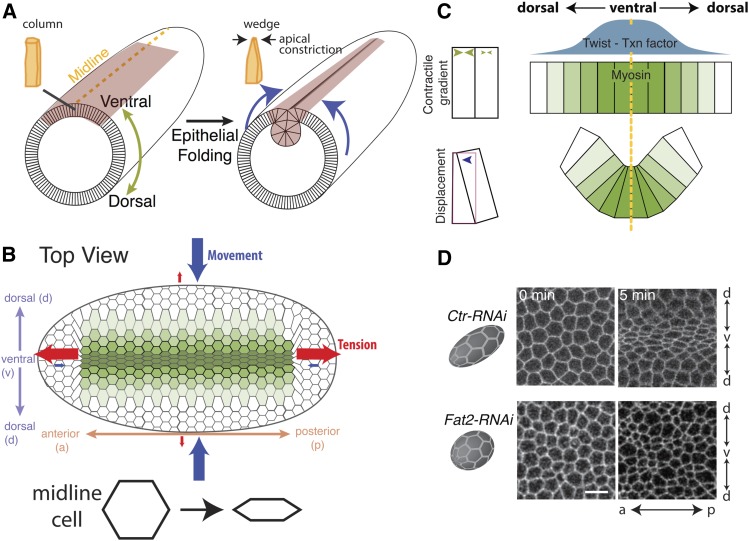
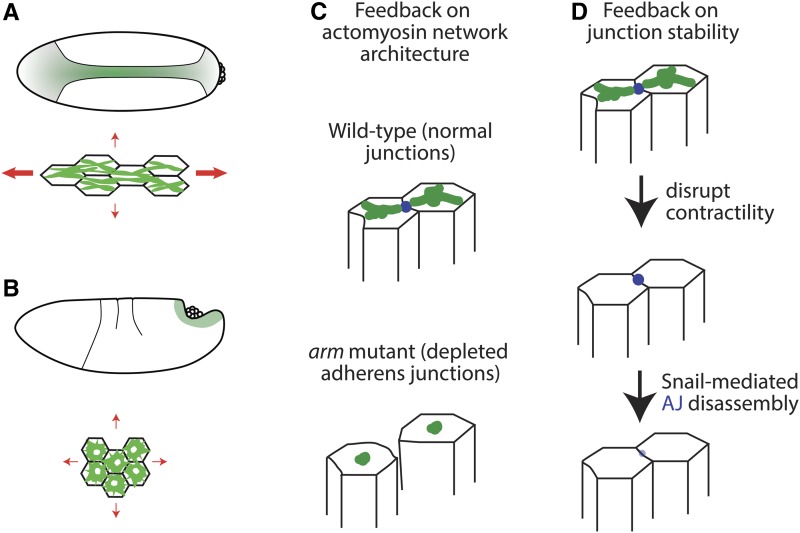
A critical juncture in early development is the partitioning of cells that will adopt different fates into three germ layers: the ectoderm, the mesoderm, and the endoderm. This step is achieved through the internalization of specified cells from the outermost surface layer, through a process called gastrulation. In , gastrulation is achieved through cell shape changes (, apical constriction) that change tissue curvature and lead to the folding of a surface epithelium. Folding of embryonic tissue results in mesoderm and endoderm invagination, not as individual cells, but as collective tissue units. The tractability of as a model system is best exemplified by how much we know about gastrulation, from the signals that pattern the embryo to the molecular components that generate force, and how these components are organized to promote cell and tissue shape changes. For mesoderm invagination, graded signaling by the morphogen, Spätzle, sets up a gradient in transcriptional activity that leads to the expression of a secreted ligand (Folded gastrulation) and a transmembrane protein (T48). Together with the GPCR Mist, which is expressed in the mesoderm, and the GPCR Smog, which is expressed uniformly, these signals activate heterotrimeric G-protein and small Rho-family G-protein signaling to promote apical contractility and changes in cell and tissue shape. A notable feature of this signaling pathway is its intricate organization in both space and time. At the cellular level, signaling components and the cytoskeleton exhibit striking polarity, not only along the apical-basal cell axis, but also within the apical domain. Furthermore, gene expression controls a highly choreographed chain of events, the dynamics of which are critical for primordium invagination; it does not simply throw the cytoskeletal "on" switch. Finally, studies of gastrulation have provided insight into how global tissue mechanics and movements are intertwined as multiple tissues simultaneously change shape. Overall, these studies have contributed to the view that cells respond to forces that propagate over great distances, demonstrating that cellular decisions, and, ultimately, tissue shape changes, proceed by integrating cues across an entire embryo.
外胚层、中胚层和内胚层。这一步是通过将特定细胞从最外层内化来实现的,这个过程称为原肠胚形成。在果蝇中,原肠胚形成是通过细胞形状变化(如顶端收缩)来实现的,这些变化改变了组织的曲率,导致表面上皮的折叠。胚胎组织的折叠导致中胚层和内胚层的内陷,不是作为单个细胞,而是作为集体组织单位。果蝇作为一个模型系统的可操作性最好地体现在我们对果蝇原肠胚形成的了解程度上,从胚胎模式形成的信号到产生力的分子成分,以及这些成分如何组织起来促进细胞和组织形状的变化。对于中胚层的内陷,形态发生素 Spätzle 的梯度信号建立了转录活性的梯度,导致分泌配体(折叠原肠胚形成)和跨膜蛋白(T48)的表达。与在中胚层中表达的 GPCR Mist 和均匀表达的 GPCR Smog 一起,这些信号激活异三聚体 G 蛋白和小 Rho 家族 G 蛋白信号,促进顶端收缩和细胞和组织形状的变化。该信号通路的一个显著特征是其在空间和时间上的精细组织。在细胞水平上,信号成分和细胞骨架表现出惊人的极性,不仅沿着顶端-基底细胞轴,而且在顶端区域内也是如此。此外,基因表达控制着高度协调的事件链,这些事件的动力学对于原基内陷至关重要;它并不是简单地打开细胞骨架“开关”。最后,对果蝇原肠胚形成的研究提供了一些见解,即多个组织同时改变形状时,全局组织力学和运动是如何相互交织的。总的来说,这些研究有助于形成这样一种观点,即细胞对传播距离很远的力做出反应,表明细胞的决策,以及最终的组织形状变化,是通过整合整个胚胎的线索来进行的。