Department of Molecular Biology and Biotechnology, Pan African University Institute for Basic Sciences, Technology & Innovation, PO Box 62000-00200, Nairobi, Kenya.
Kenya Agricultural and Livestock Research Organization, Biotechnology Research Institute, PO Box 362-00902, Kikuyu, Kenya.
Parasit Vectors. 2020 May 14;13(1):253. doi: 10.1186/s13071-020-04120-3.
Despite the morphological characterization established in the 1950s and 1960s, the identity of extant taxa that make up Glossina fuscipes (s.l.) in the Congo remains questionable. Previous claims of overlap between G. fuscipes (believed to be G. f. quanzensis) and G. palpalis palpalis around Brazzaville city further complicate the taxonomic status and population dynamics of the two taxa. This study aimed to determine the phylogenetic relationships between G. fuscipes (s.l.) and G. p. palpalis and to assess genetic variation among G. fuscipes (s.l.) populations in Congo Brazzaville.
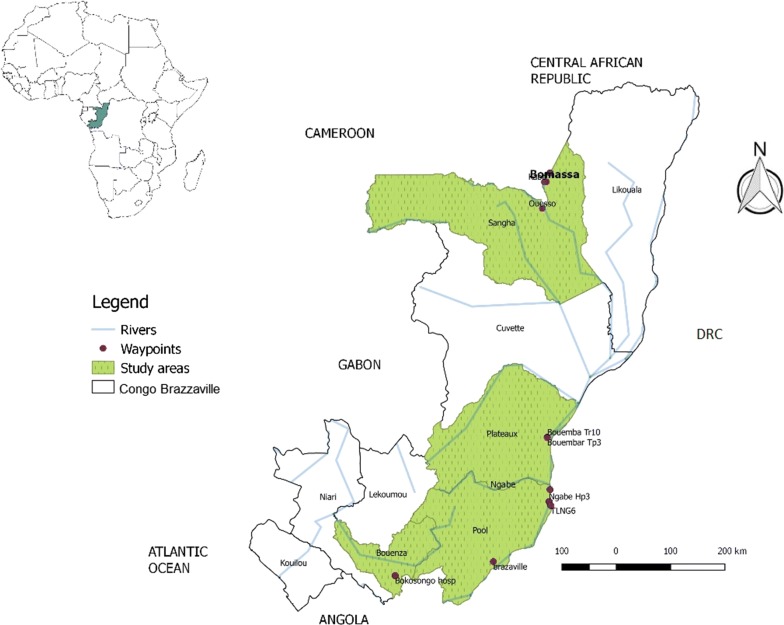
We collected 263 G. fuscipes (s.l.) from northern and central regions, and 65 G. p. palpalis from southern part of the country. The mitochondrial cytochrome c oxidase subunit 1 (cox1) gene was amplified using taxa-specific primer pairs. Sequence data were analyzed in DnaSP and Arlequin to assess the genetic diversity, differentiation and demographic history of G. fuscipes (s.l.) populations.
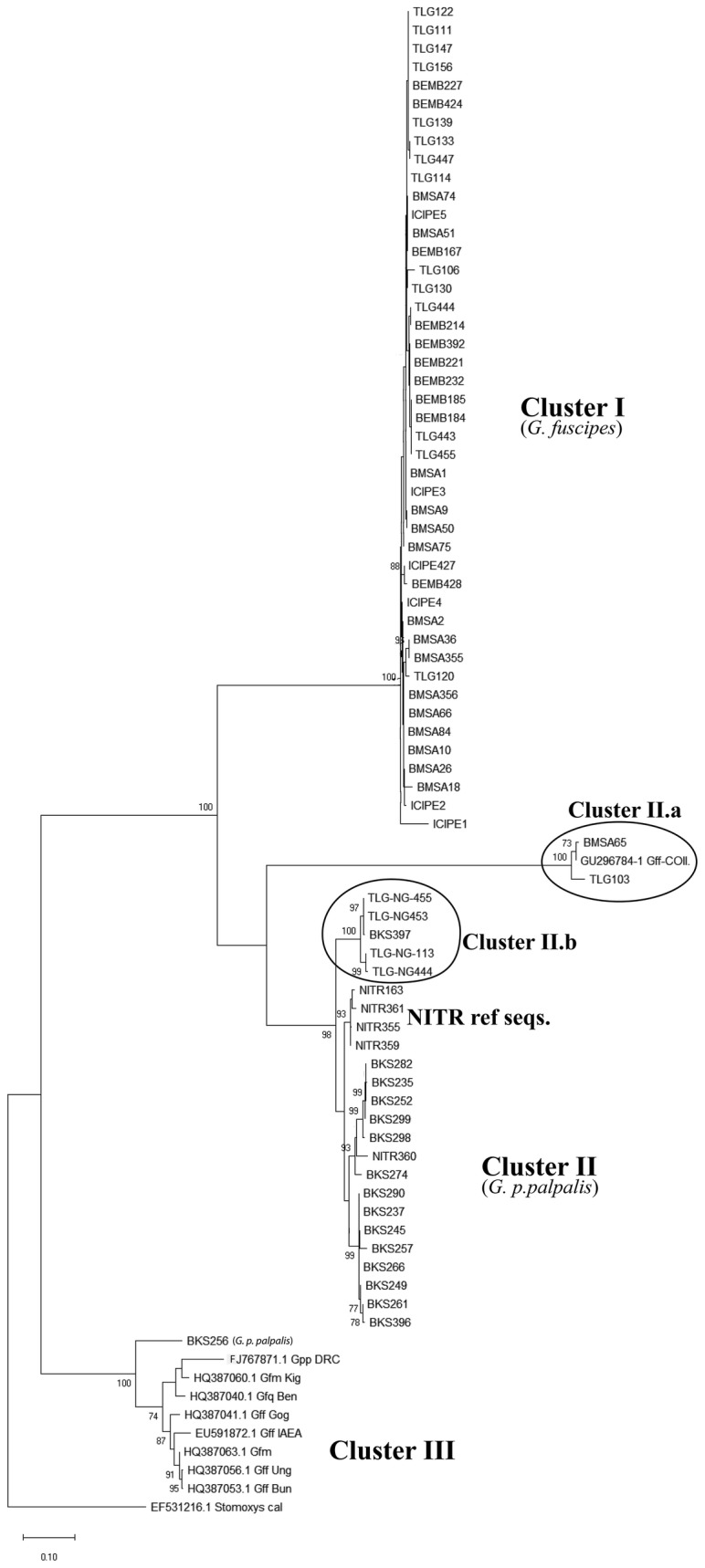
The general BLAST analysis yielded a similarity of 99% for G. fuscipes (s.l.) and G. p. palpalis. BLASTn analysis for G. fuscipes (s.l.) showed > 98% identity with GenBank sequences for G. fuscipes (s.l.), with BEMB population showing 100% similarity with G. f. fuscipes. Glossina fuscipes (s.l.) populations showed high haplotype diversity (H = 46, Hd = 0.884), moderate nucleotide diversity ( = 0.012) and moderate (F = 0.072) to high (F = 0.152) genetic differentiation. Most of the genetic variation (89.73%) was maintained within populations. The mismatch analysis and neutrality tests indicated recent tsetse population expansions.
Phylogenetic analysis revealed minor differences between G. fuscipes (s.l.) and G. p. palpalis. Genetic diversity of G. fuscipes (s.l.) was high in the populations sampled except one. Genetic differentiation ranged from moderate to high among subpopulations. There was a restricted gene flow between G. fuscipes (s.l.) populations in the north and central part of the country. Genetic signatures based on cox1 showed recent expansion and recovery of G. fuscipes (s.l.) populations from previous bottlenecks. To fully understand the species distribution limits, we recommend further studies involving a wider sampling scheme including the swampy Mossaka focus for G. fuscipes (s.l.) and the entire range of G. p. palpalis in South Congo.
尽管在 20 世纪 50 年代和 60 年代已经确定了形态特征,但构成刚果金沙蝇(s.l.)的现存分类群的身份仍然存在疑问。此前有报道称,金沙蝇(被认为是 G. f. quanzensis)与布拉萨市周围的 G. palpalis palpalis 之间存在重叠,这进一步使这两个分类群的分类地位和种群动态复杂化。本研究旨在确定金沙蝇(s.l.)与 G. palpalis palpalis 之间的系统发育关系,并评估金沙蝇(s.l.)在刚果布拉柴维尔的种群遗传变异。
我们从北部和中部地区收集了 263 只金沙蝇(s.l.),从该国南部收集了 65 只 G. p. palpalis。使用种特异性引物对扩增线粒体细胞色素 c 氧化酶亚基 1(cox1)基因。在 DnaSP 和 Arlequin 中分析序列数据,以评估金沙蝇(s.l.)种群的遗传多样性、分化和种群历史。
普通 BLAST 分析显示金沙蝇(s.l.)和 G. p. palpalis 的相似度为 99%。金沙蝇(s.l.)的 BLASTn 分析显示,与 GenBank 序列的相似度 > 98%,BEMB 种群与 G. f. fuscipes 完全相同。金沙蝇(s.l.)种群表现出高单倍型多样性(H = 46,Hd = 0.884)、中等核苷酸多样性( = 0.012)和中等(F = 0.072)到高(F = 0.152)的遗传分化。大部分遗传变异(89.73%)存在于种群内。错配分析和中性检验表明,最近有舌蝇种群扩张。
系统发育分析显示金沙蝇(s.l.)和 G. p. palpalis 之间存在细微差异。除一个种群外,所采样的金沙蝇(s.l.)种群遗传多样性较高。亚种群之间的遗传分化程度从中等到高度不等。该国北部和中部的金沙蝇(s.l.)种群之间存在有限的基因流。基于 cox1 的遗传特征表明,金沙蝇(s.l.)种群经历了最近的扩张和从以前的瓶颈中恢复。为了充分了解物种分布范围,我们建议进一步开展研究,包括扩大采样范围,包括 Mossaka 沼泽地的金沙蝇(s.l.)和南刚果的 G. p. palpalis 整个范围。