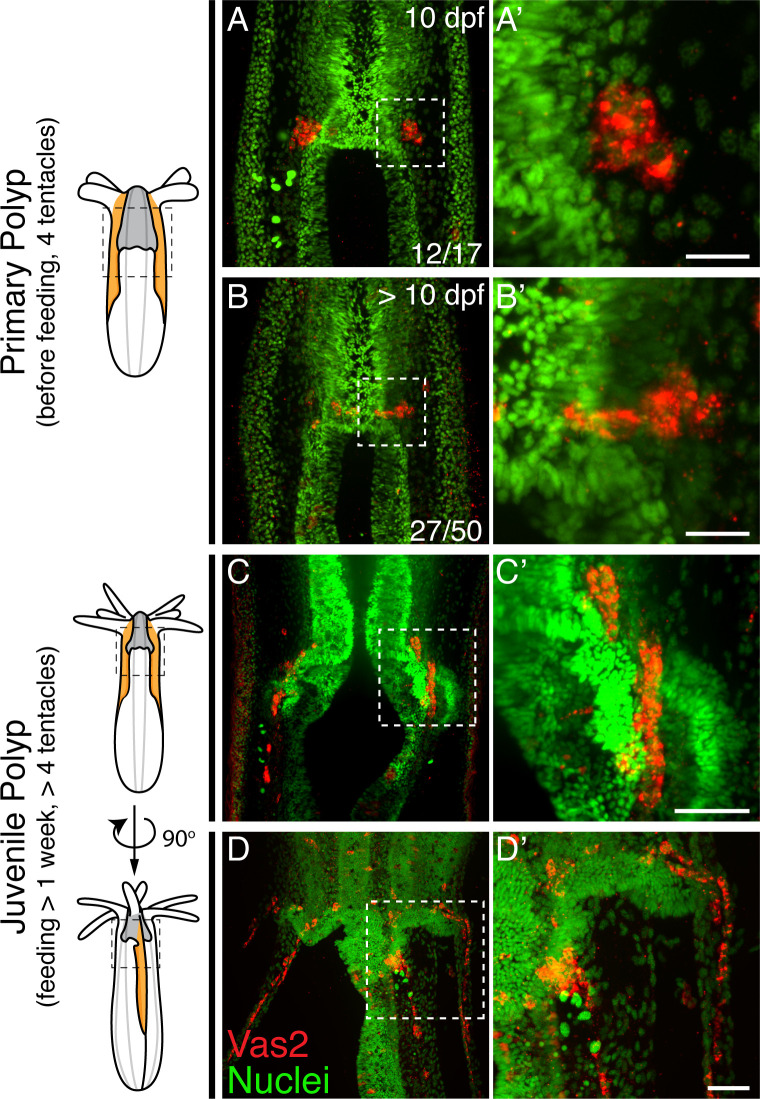
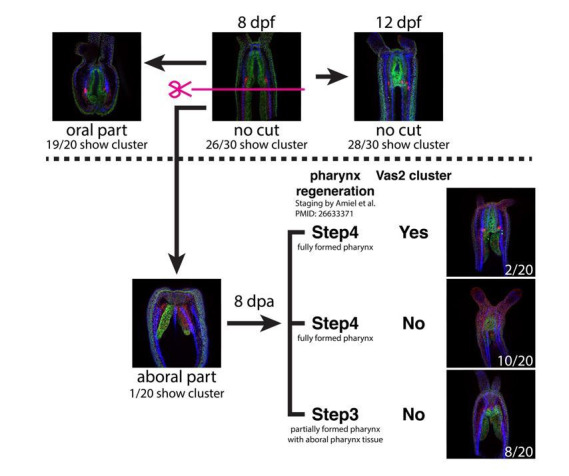
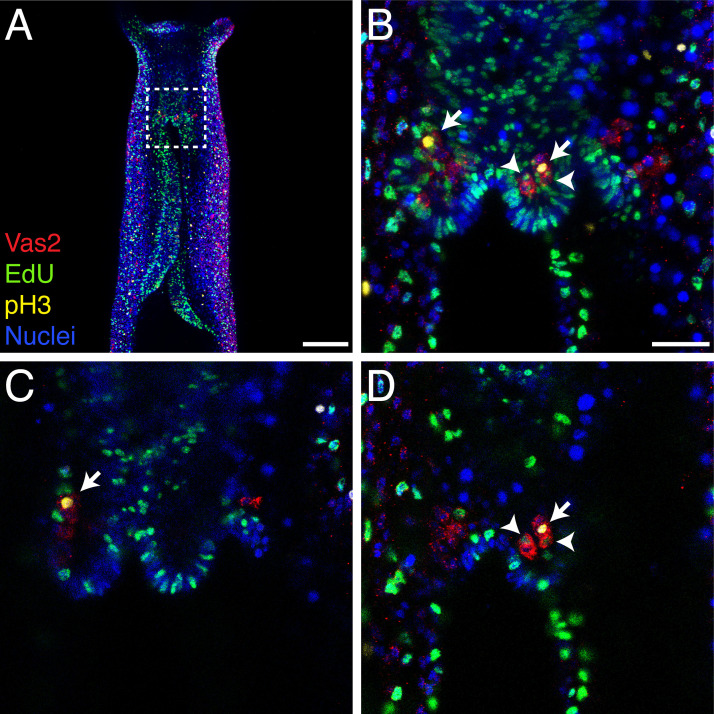
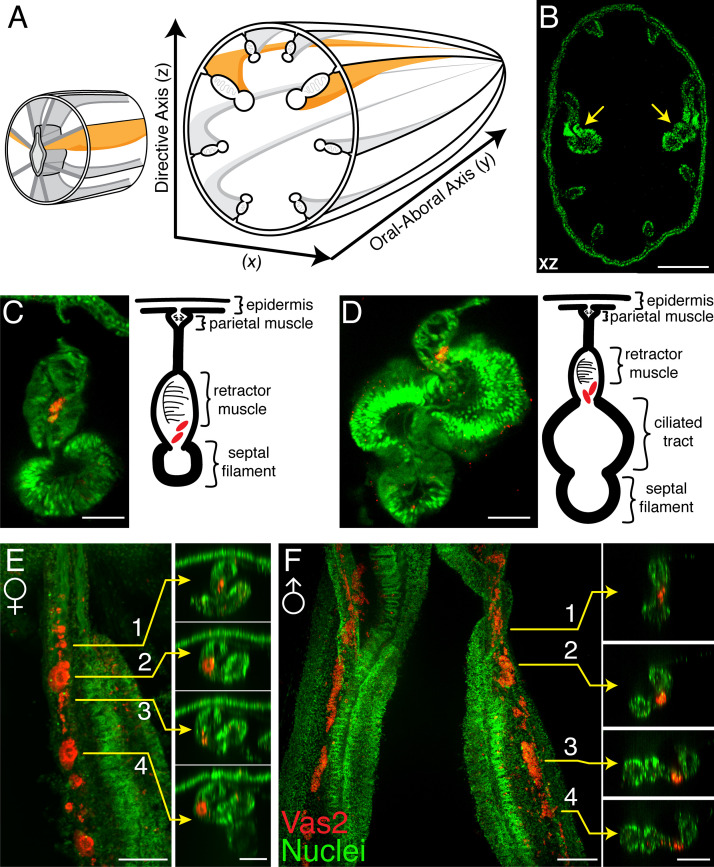
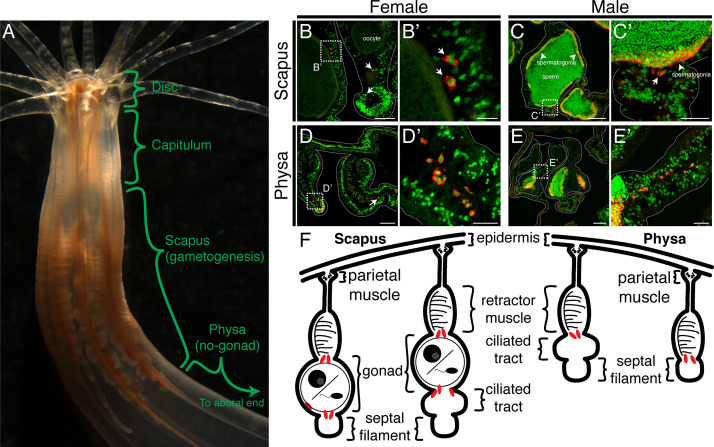
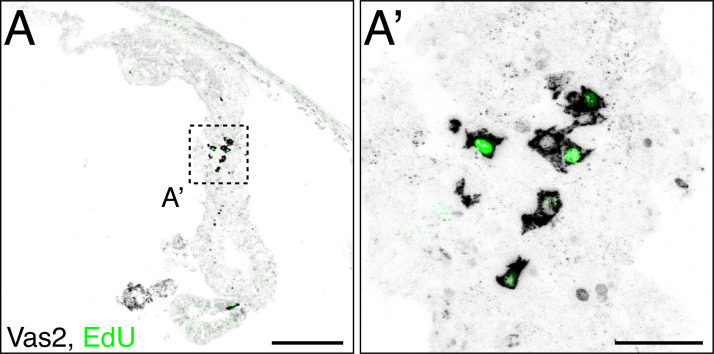
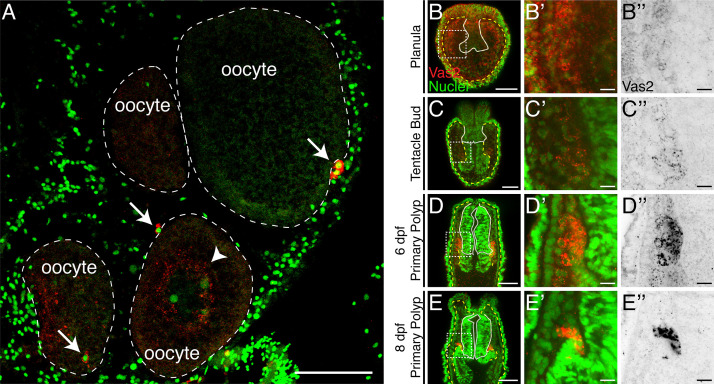
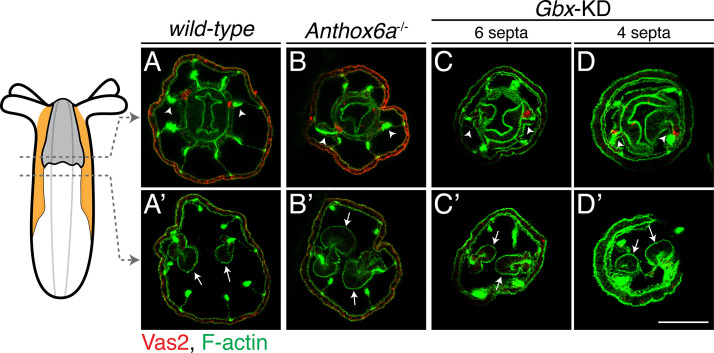
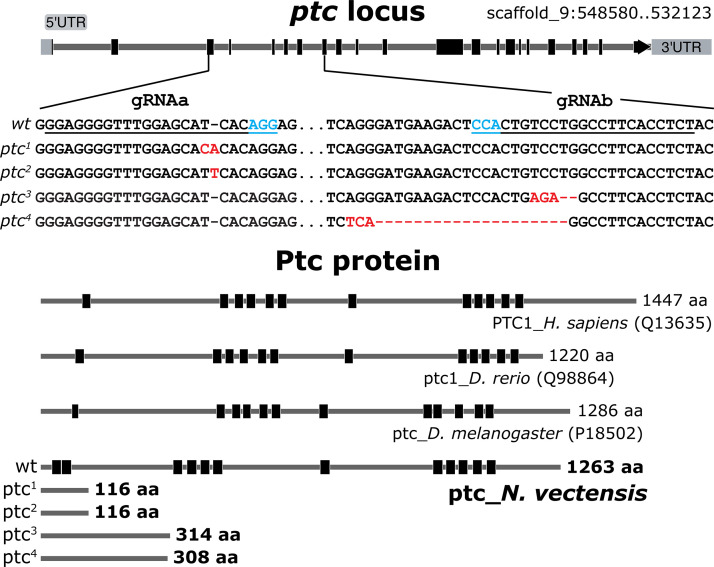
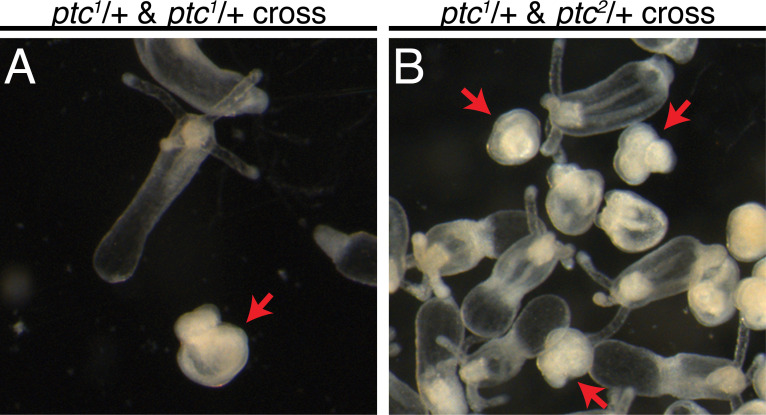
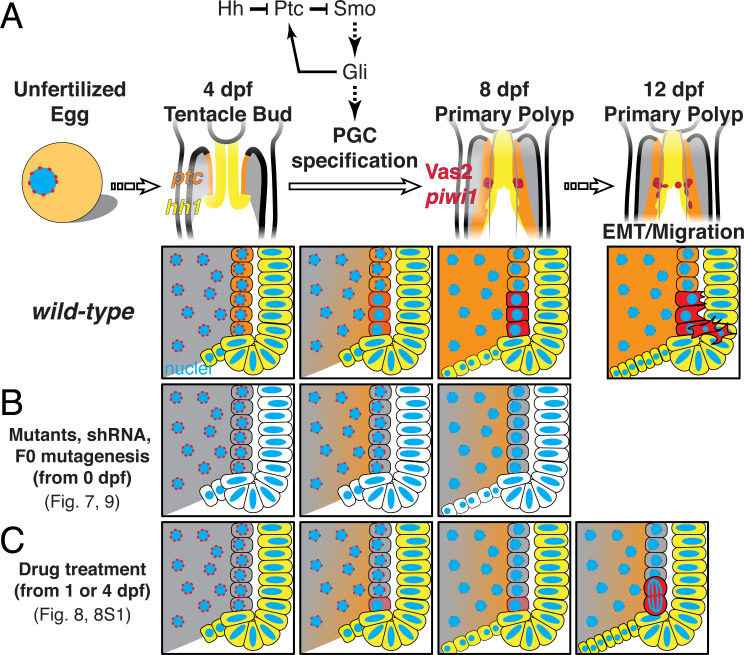
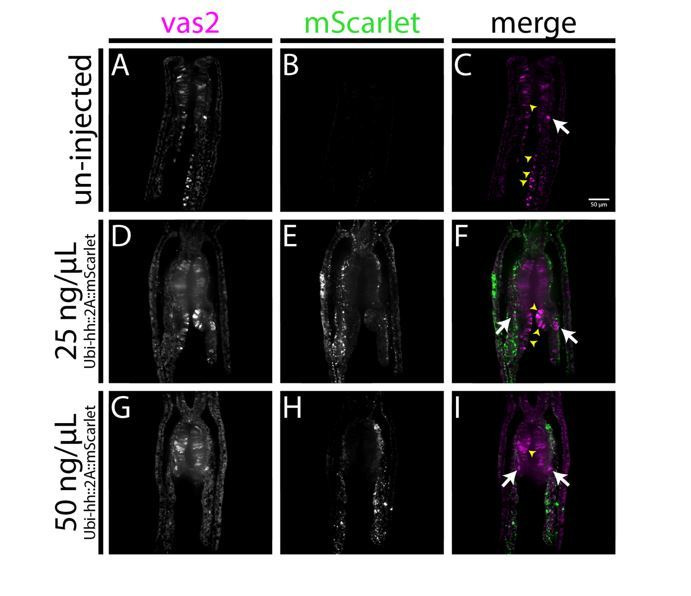
Stowers Institute for Medical Research, Kansas City, United States.
Department of Anatomy and Cell Biology, The University of Kansas School of Medicine, Kansas City, United States.
Elife. 2020 Sep 24;9:e54573. doi: 10.7554/eLife.54573.
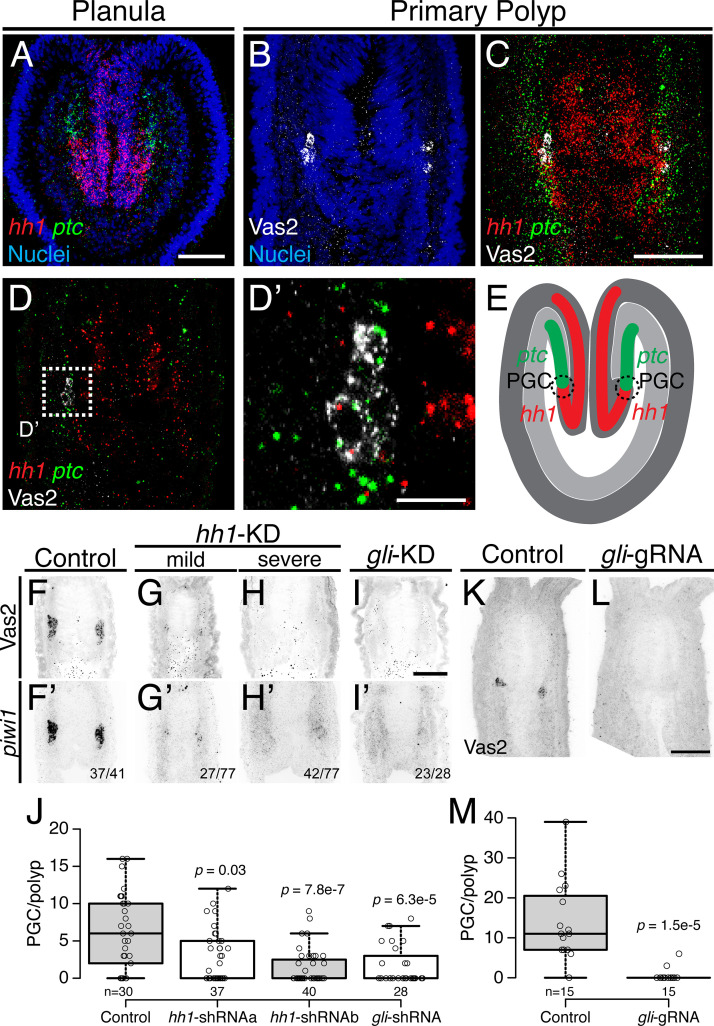
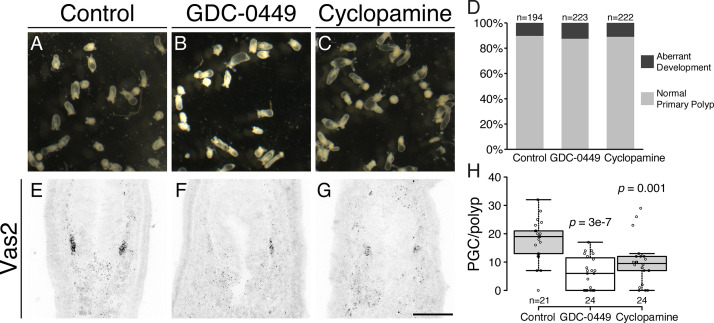
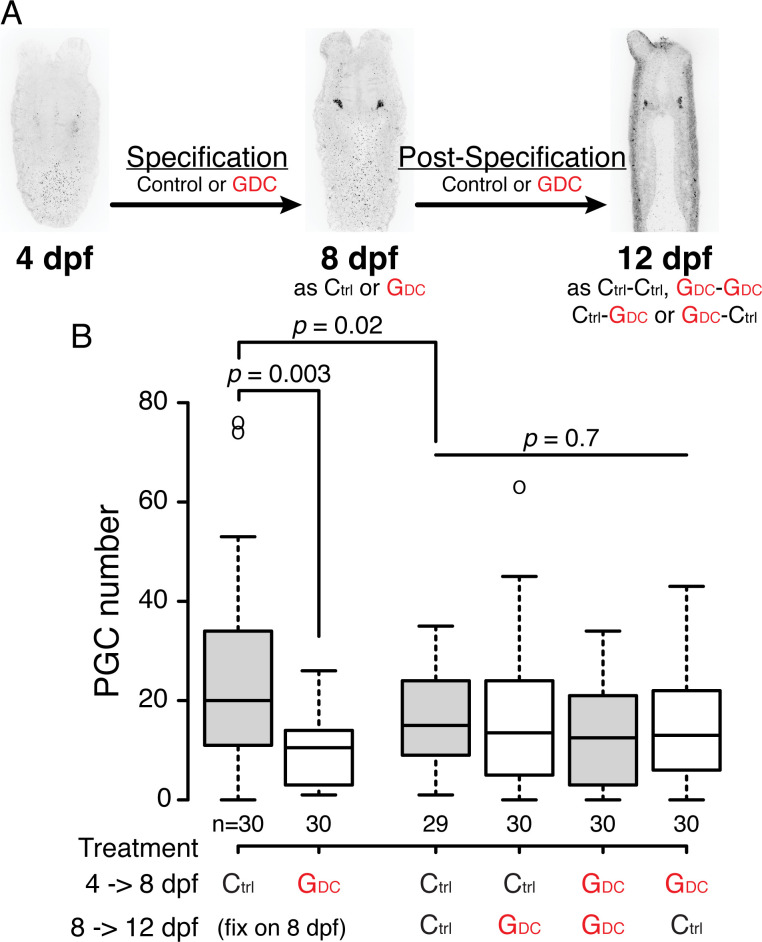
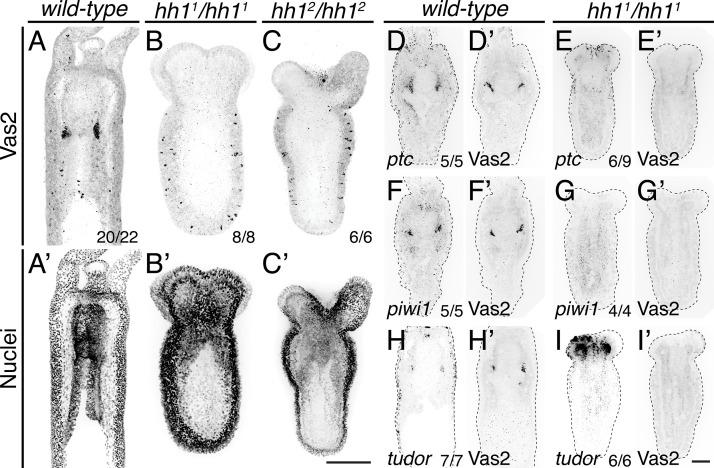
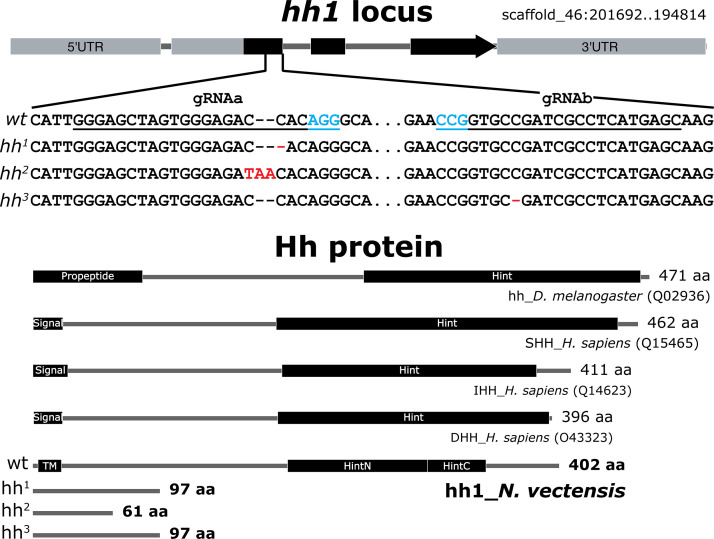
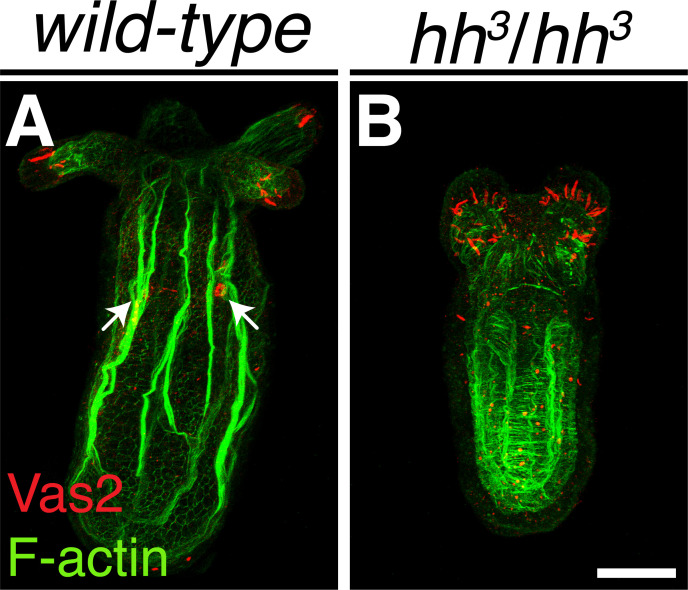
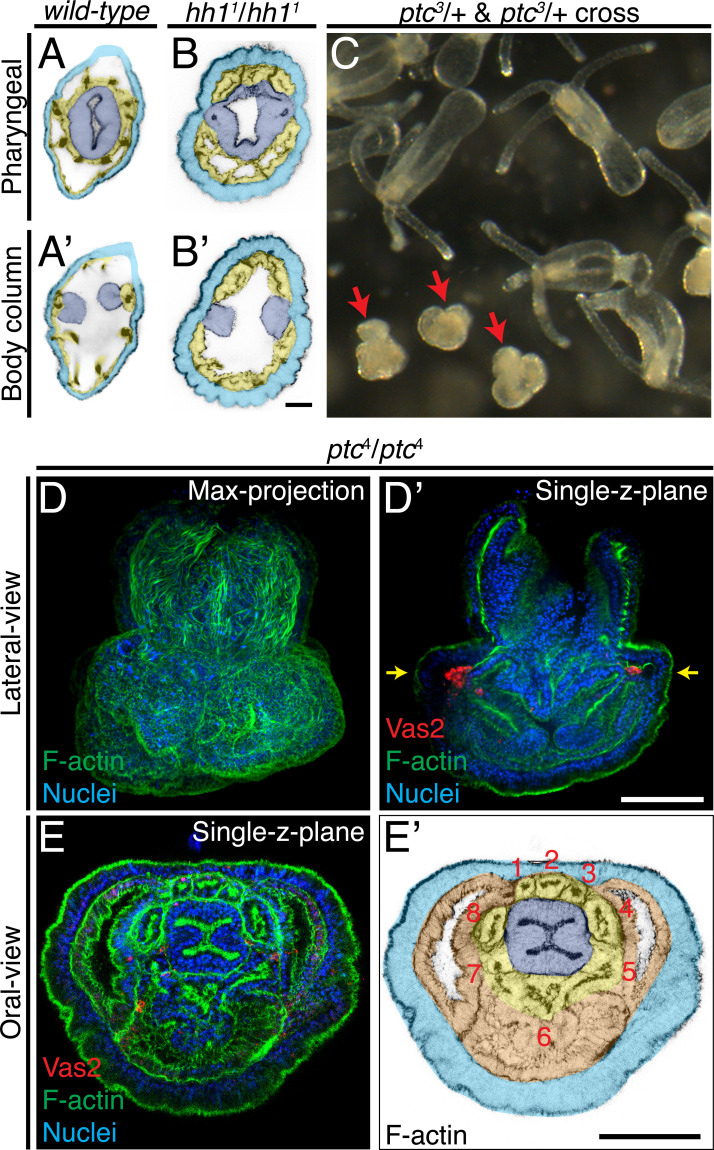
Two distinct mechanisms for primordial germ cell (PGC) specification are observed within Bilatera: early determination by maternal factors or late induction by zygotic cues. Here we investigate the molecular basis for PGC specification in , a representative pre-bilaterian animal where PGCs arise as paired endomesodermal cell clusters during early development. We first present evidence that the putative PGCs delaminate from the endomesoderm upon feeding, migrate into the gonad primordia, and mature into germ cells. We then show that the PGC clusters arise at the interface between and domains in the developing mesenteries and use gene knockdown, knockout and inhibitor experiments to demonstrate that Hh signaling is required for both PGC specification and general endomesodermal patterning. These results provide evidence that the germline is specified by inductive signals rather than maternal factors, and support the existence of zygotically-induced PGCs in the eumetazoan common ancestor.
在两侧对称动物中观察到两种截然不同的原始生殖细胞(PGC)特化机制:早期由母体因子决定或晚期由合子线索诱导。在这里,我们研究了 PGC 特化的分子基础,这是一种有代表性的前两侧对称动物,其中 PGC 在早期发育过程中作为成对的内胚层和中胚层细胞簇出现。我们首先提出证据表明,PGC 从内胚层和中胚层脱离后在进食时迁移到性腺原基中,并成熟为生殖细胞。然后我们表明,PGC 簇出现在正在发育的肠系膜的 和 区域之间的界面上,并使用基因敲低、敲除和抑制剂实验证明 Hh 信号对于 PGC 特化和一般内胚层中胚层模式形成都是必需的。这些结果提供了证据表明,生殖系是由诱导信号而不是母体因子决定的,并且支持在后生动物的共同祖先中存在合子诱导的 PGC。