National Centre for Biological Sciences, Tata Institute of Fundamental Research, Bangalore, Karnataka, India.
School of Life Science, The University of Trans-Disciplinary Health Sciences & Technology (TDU), Bangalore, Karnataka, India.
Genome Biol Evol. 2020 Dec 6;12(12):2450-2466. doi: 10.1093/gbe/evaa223.
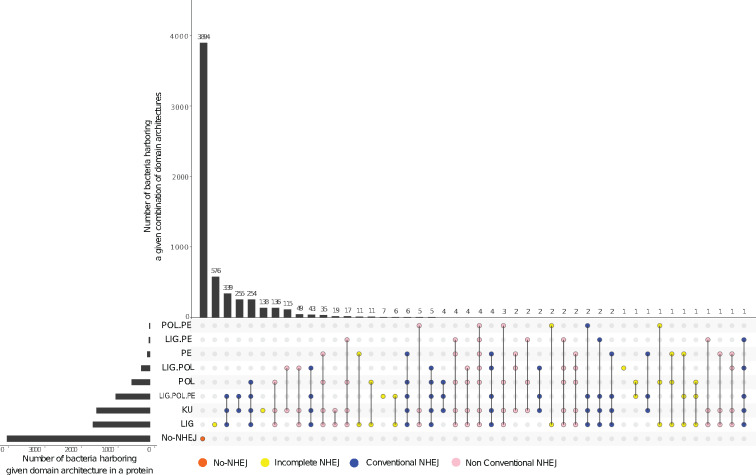
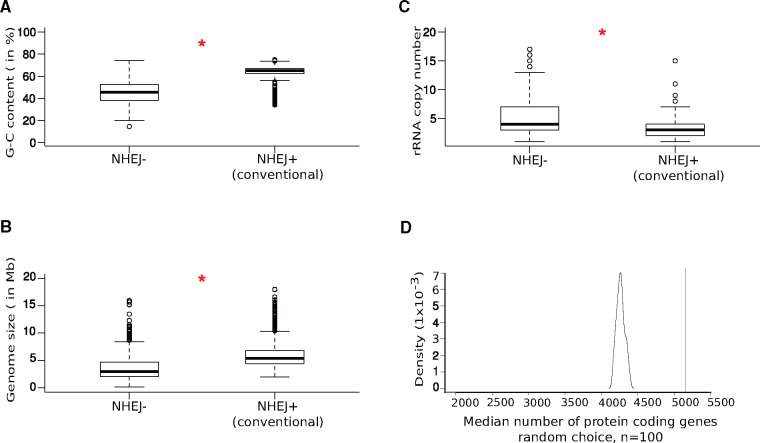
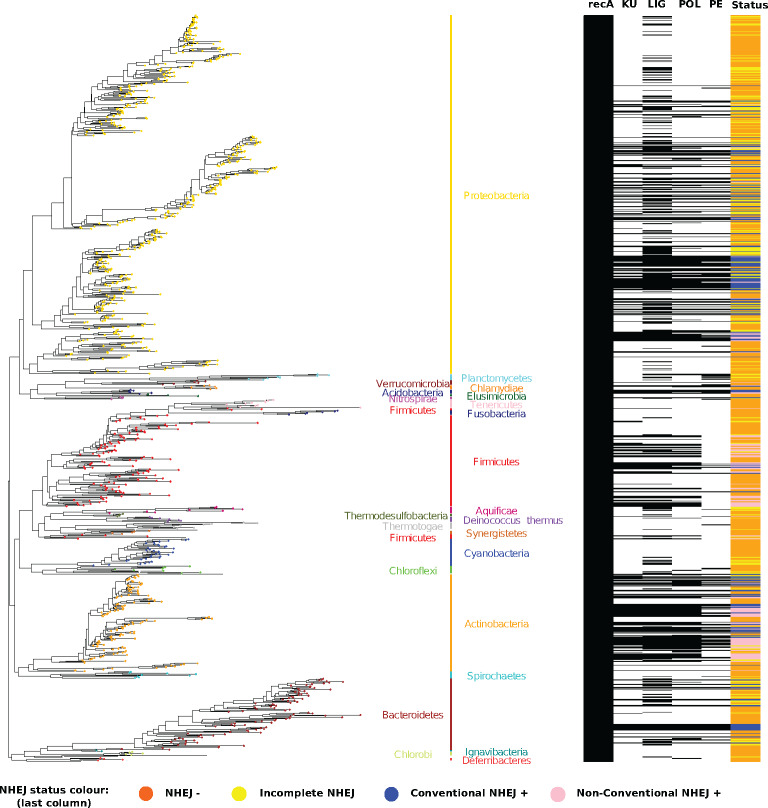
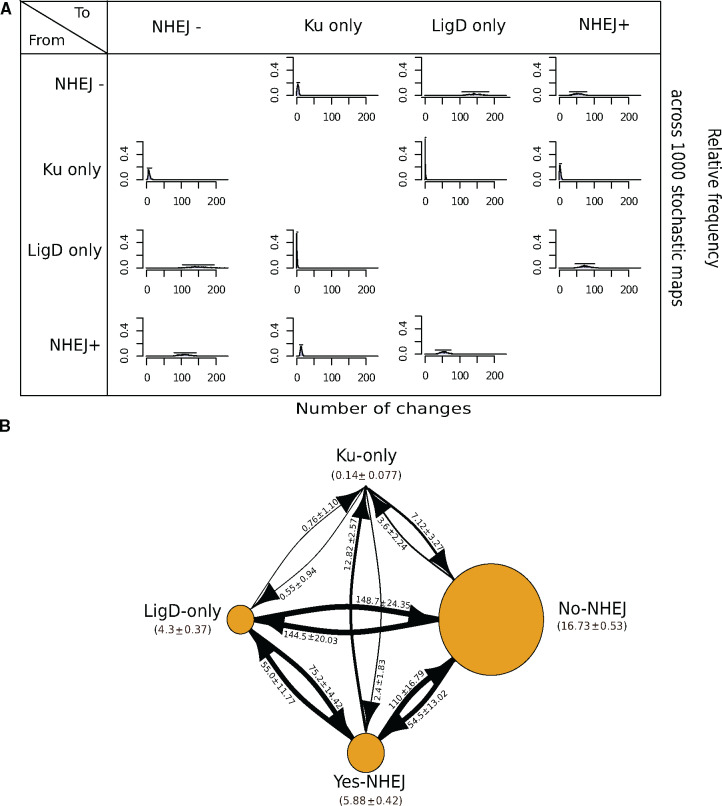
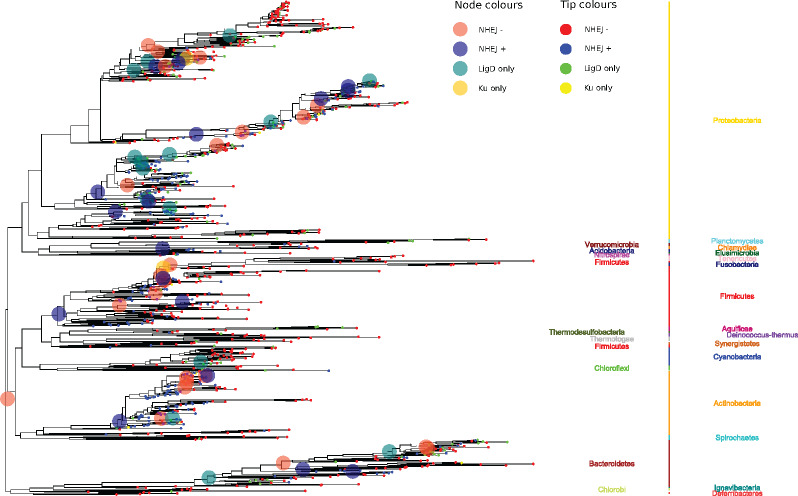
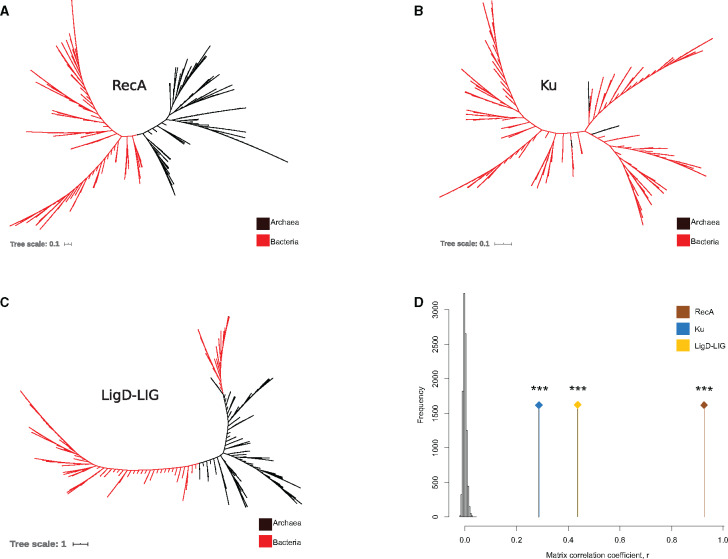
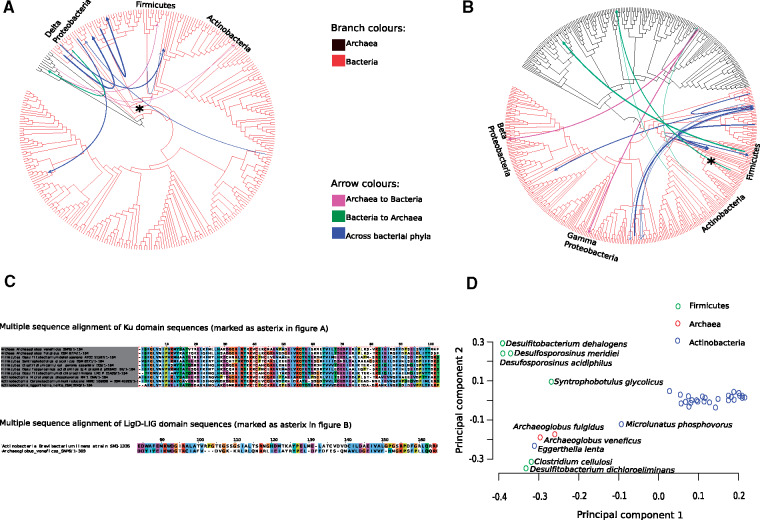
DNA double-strand breaks (DSBs) are a threat to genome stability. In all domains of life, DSBs are faithfully fixed via homologous recombination. Recombination requires the presence of an uncut copy of duplex DNA which is used as a template for repair. Alternatively, in the absence of a template, cells utilize error-prone nonhomologous end joining (NHEJ). Although ubiquitously found in eukaryotes, NHEJ is not universally present in bacteria. It is unclear as to why many prokaryotes lack this pathway. Toward understanding what could have led to the current distribution of bacterial NHEJ, we carried out comparative genomics and phylogenetic analysis across ∼6,000 genomes. Our results show that this pathway is sporadically distributed across the phylogeny. Ancestral reconstruction further suggests that NHEJ was absent in the eubacterial ancestor and can be acquired via specific routes. Integrating NHEJ occurrence data for archaea, we also find evidence for extensive horizontal exchange of NHEJ genes between the two kingdoms as well as across bacterial clades. The pattern of occurrence in bacteria is consistent with correlated evolution of NHEJ with key genome characteristics of genome size and growth rate; NHEJ presence is associated with large genome sizes and/or slow growth rates, with the former being the dominant correlate. Given the central role these traits play in determining the ability to carry out recombination, it is possible that the evolutionary history of bacterial NHEJ may have been shaped by requirement for efficient DSB repair.
DNA 双链断裂 (DSBs) 对基因组稳定性构成威胁。在所有生命领域中,DSBs 都通过同源重组被准确地修复。重组需要存在未切割的双链 DNA 拷贝,该拷贝被用作修复的模板。或者,在没有模板的情况下,细胞利用易错的非同源末端连接 (NHEJ)。虽然 NHEJ 在真核生物中普遍存在,但在细菌中并非普遍存在。目前尚不清楚为什么许多原核生物缺乏这种途径。为了了解是什么导致了目前细菌 NHEJ 的分布,我们对大约 6000 个基因组进行了比较基因组学和系统发育分析。我们的研究结果表明,该途径在系统发育中呈散在分布。祖先重建进一步表明,NHEJ 在真细菌祖先中不存在,可以通过特定途径获得。整合古菌的 NHEJ 发生数据,我们还发现了证据表明,NHEJ 基因在两个王国之间以及在细菌进化枝之间存在广泛的水平交换。细菌中发生的模式与 NHEJ 与基因组大小和生长速率等关键基因组特征的相关性进化一致;NHEJ 的存在与大基因组大小和/或慢生长速率相关,前者是主要的相关因素。鉴于这些特征在决定重组能力方面的核心作用,细菌 NHEJ 的进化历史可能受到有效 DSB 修复的需求所塑造。