Neurosciences and Cellular and Structural Biology Division, Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892, USA.
Neurosciences and Cellular and Structural Biology Division, Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892, USA.
Curr Biol. 2021 Feb 8;31(3):540-554.e5. doi: 10.1016/j.cub.2020.10.071. Epub 2020 Nov 23.
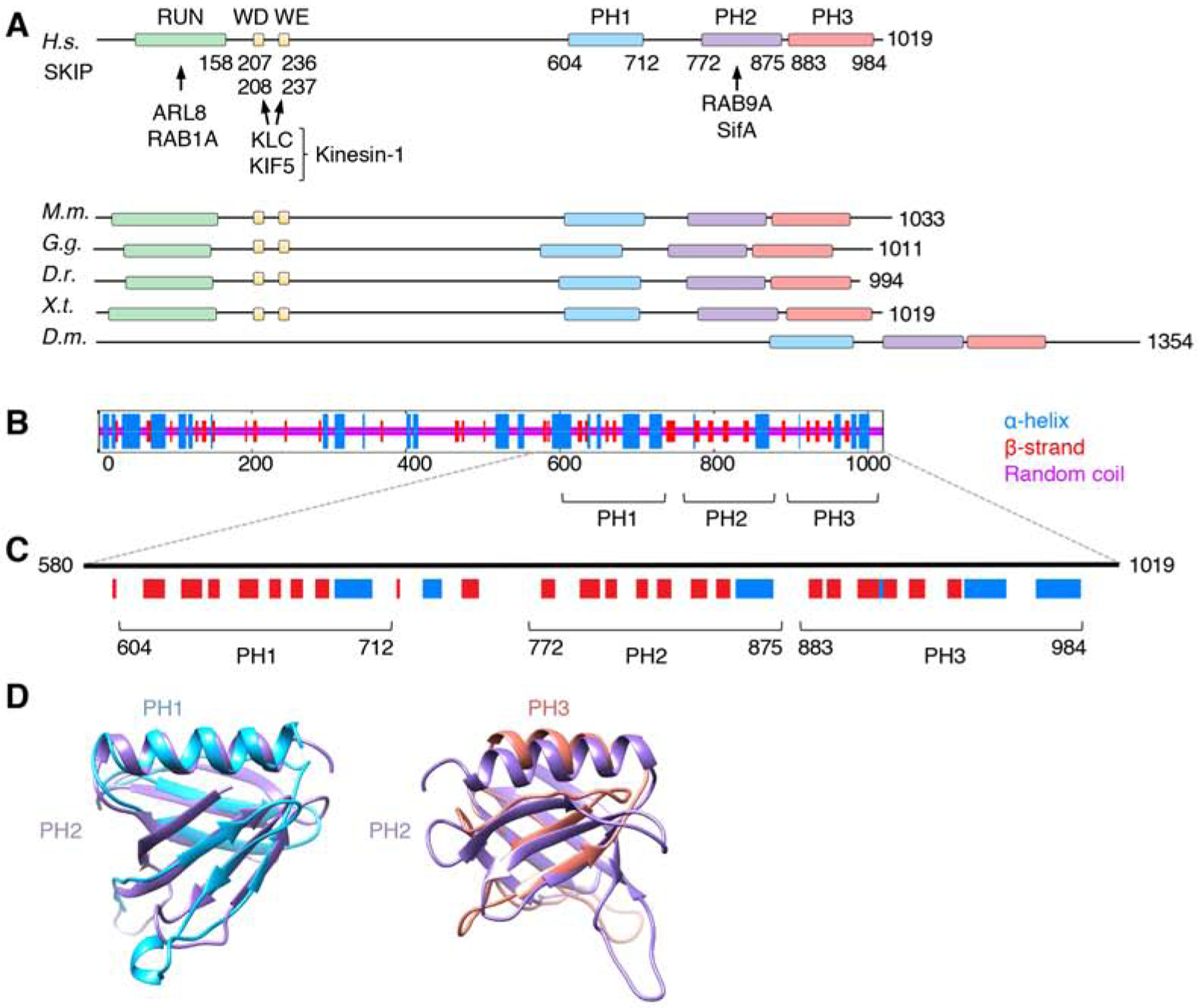
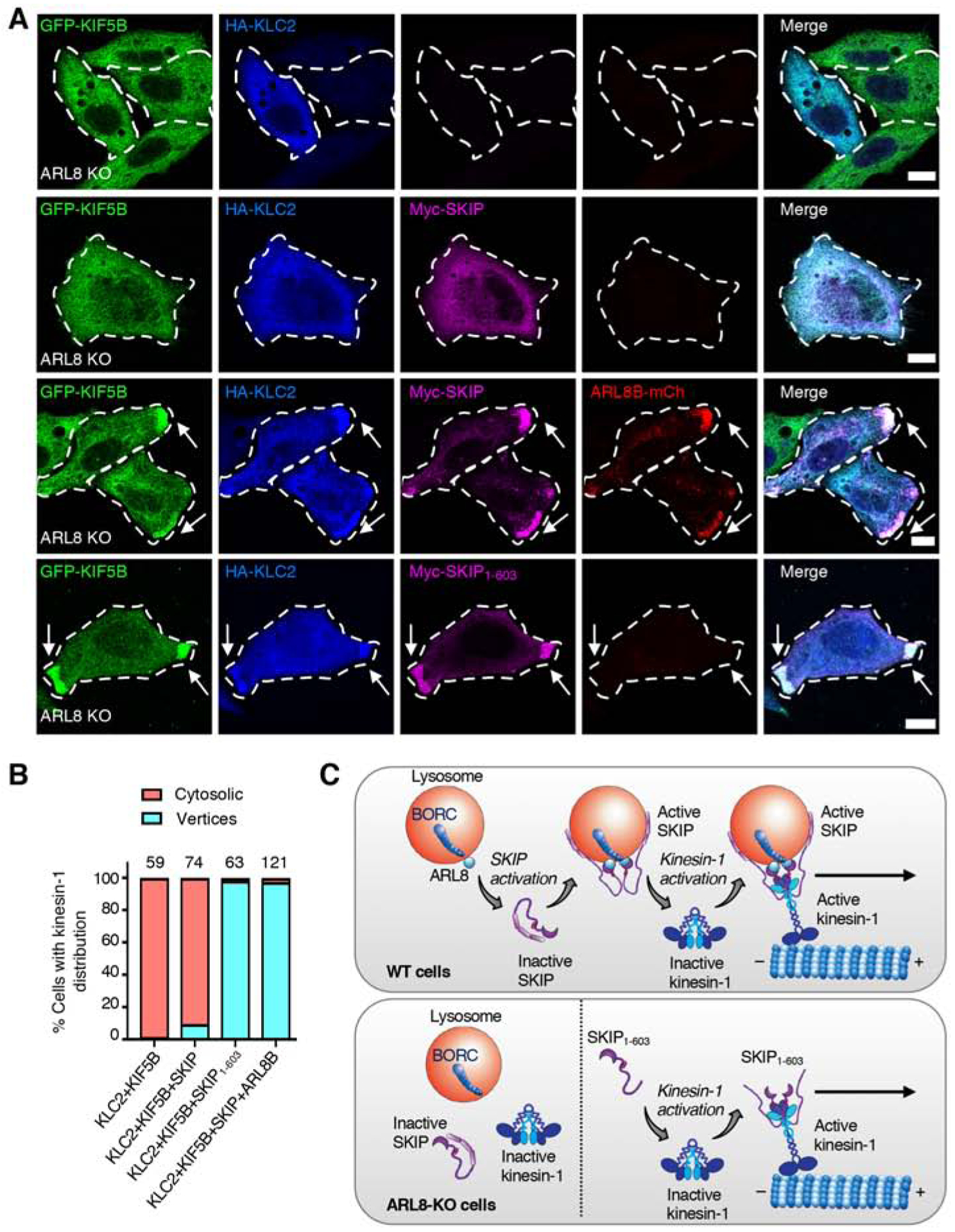
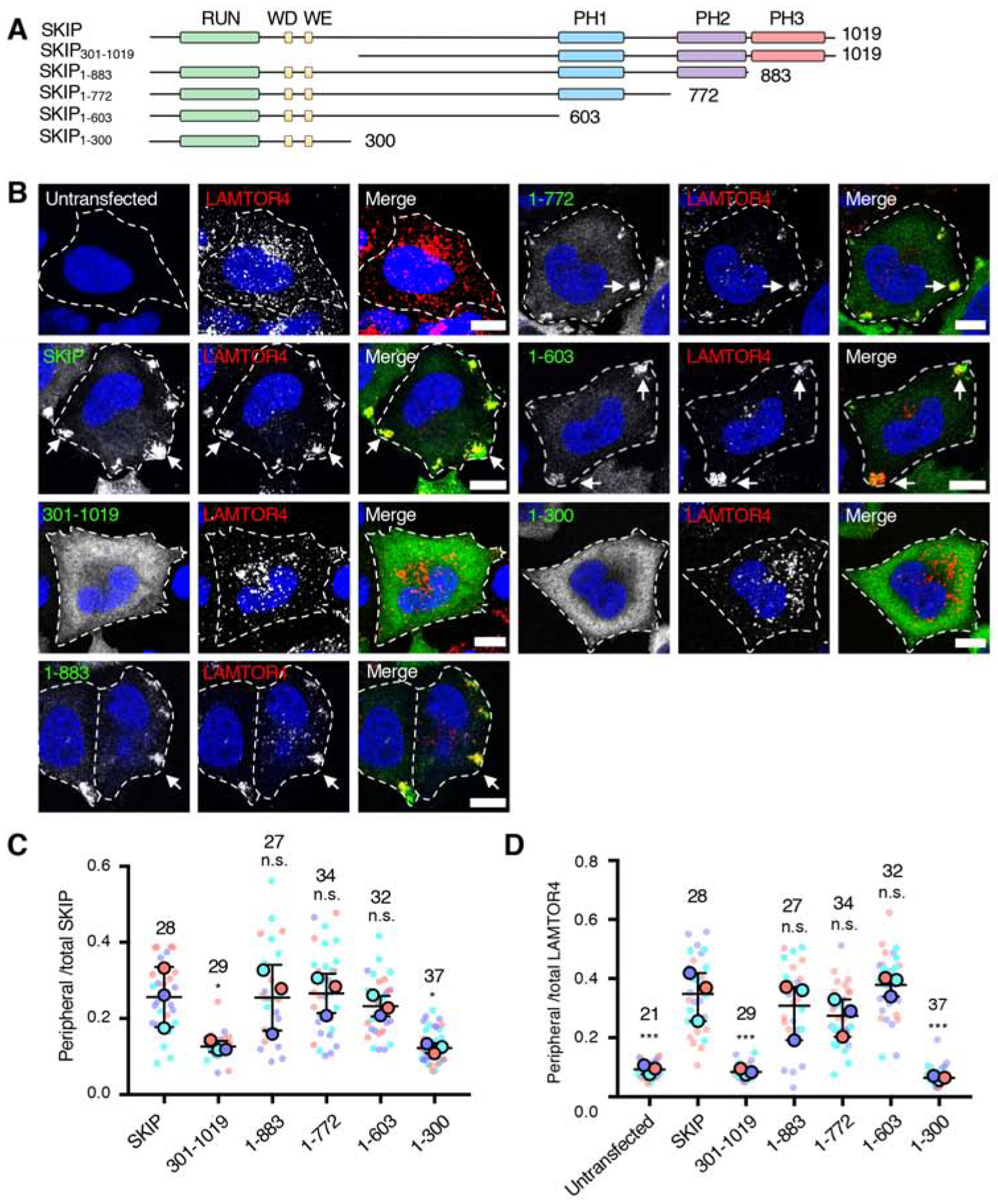
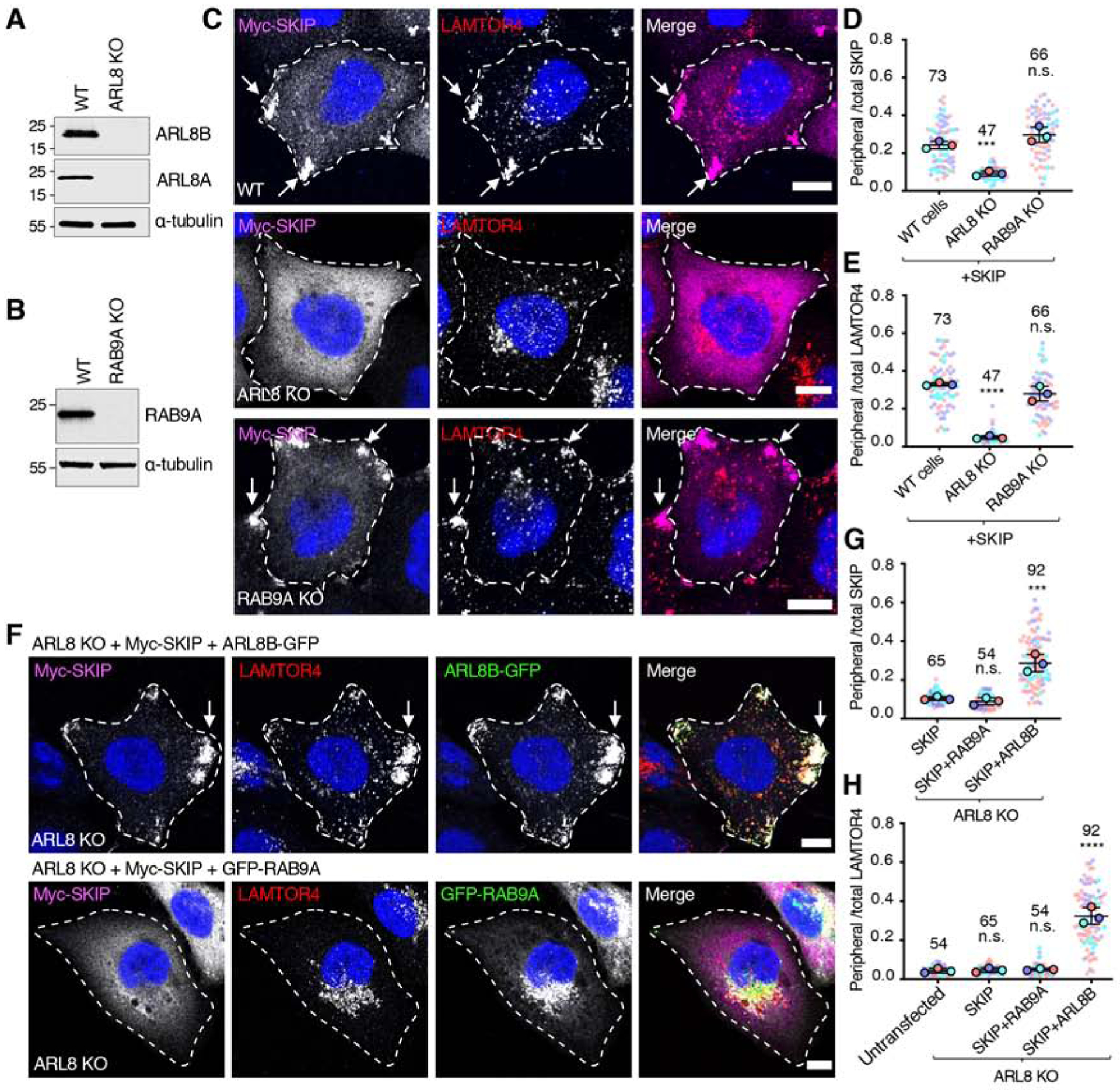
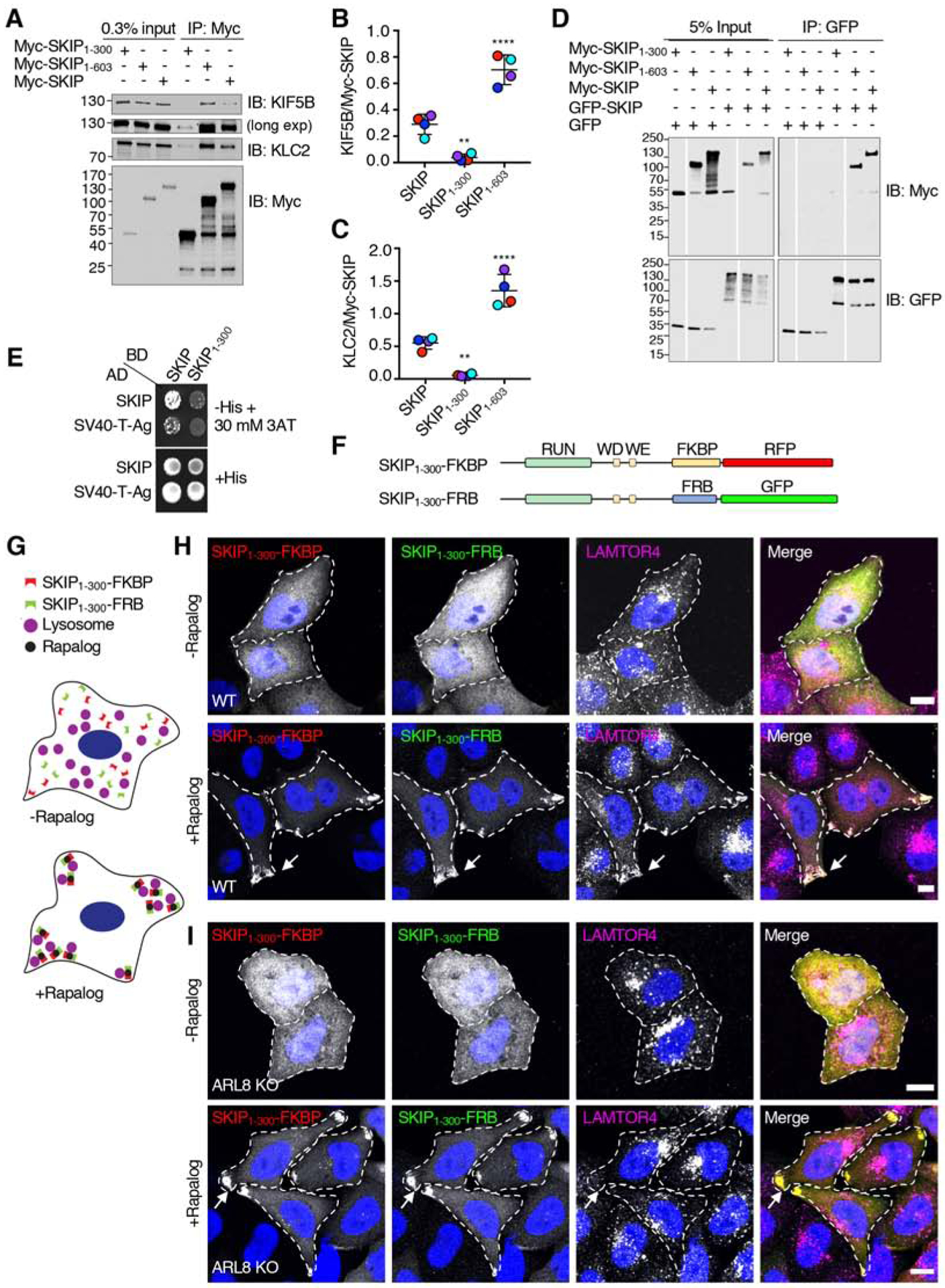
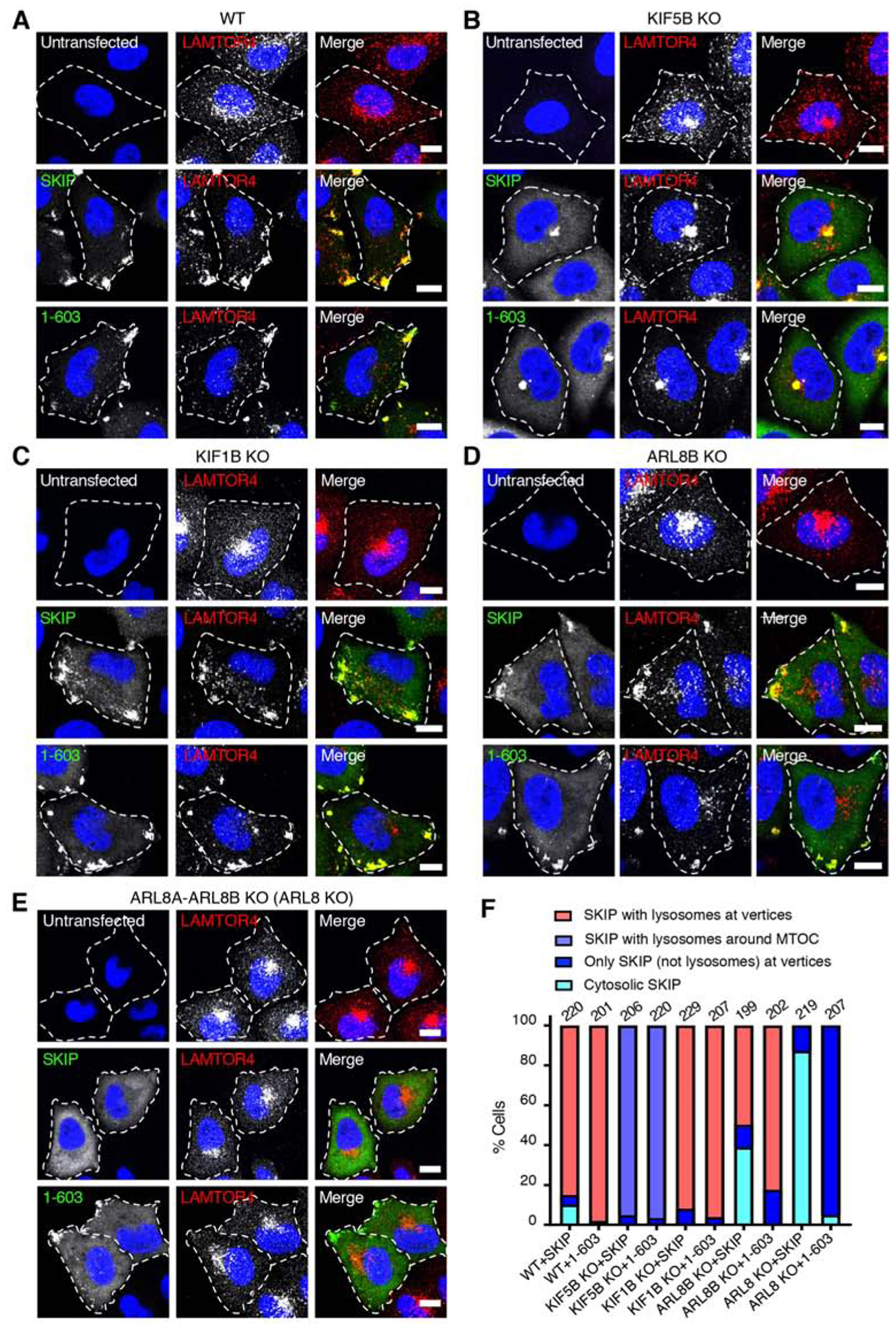
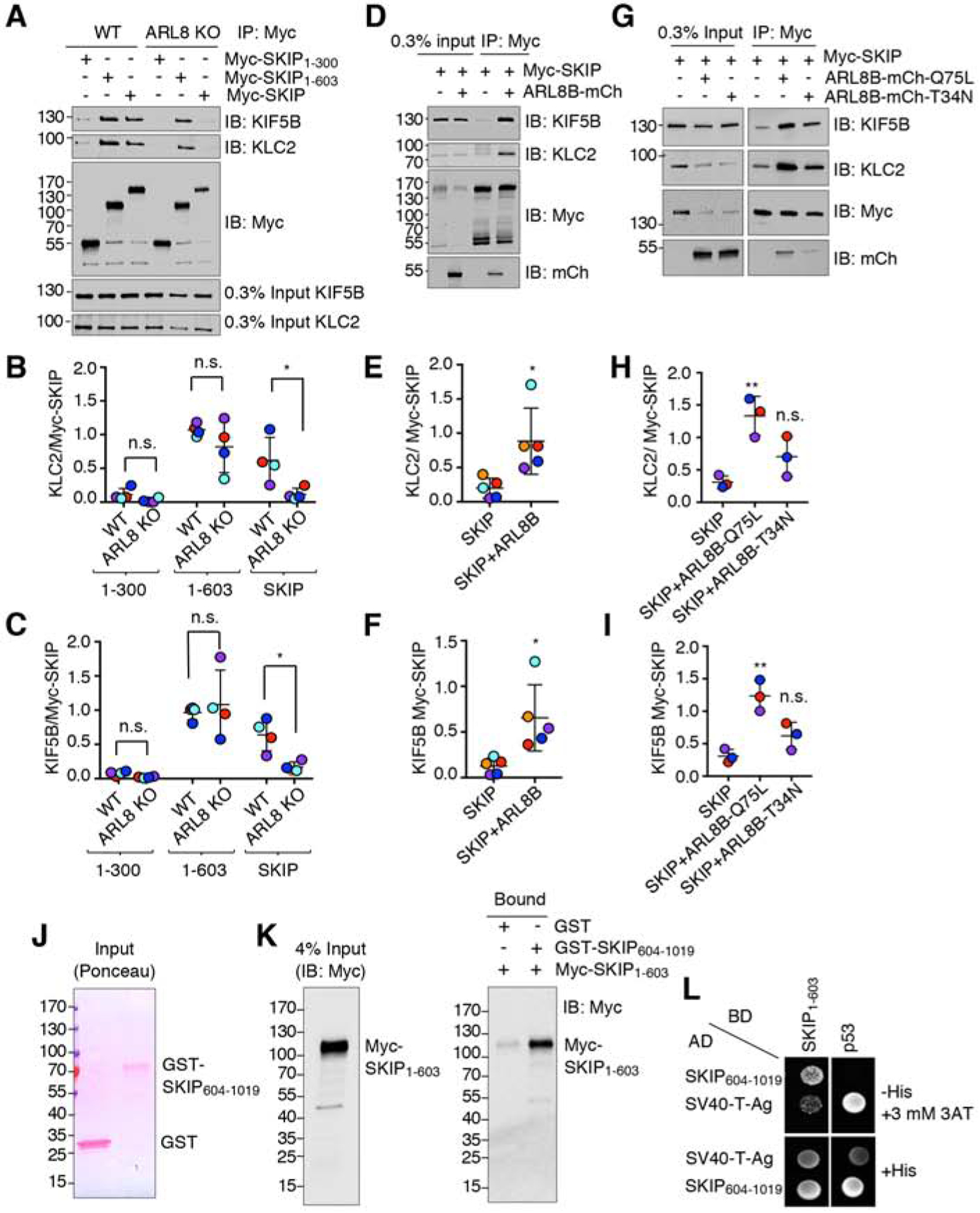
Long-range movement of organelles within the cytoplasm relies on coupling to microtubule motors, a process that is often mediated by adaptor proteins. In many cases, this coupling involves organelle- or adaptor-induced activation of the microtubule motors by conformational reversal of an autoinhibited state. Herein, we show that a similar regulatory mechanism operates for an adaptor protein named SKIP (also known as PLEKHM2). SKIP binds to the small guanosine triphosphatase (GTPase) ARL8 on the lysosomal membrane to couple lysosomes to the anterograde microtubule motor kinesin-1. Structure-function analyses of SKIP reveal that the C-terminal region comprising three pleckstrin homology (PH) domains interacts with the N-terminal region comprising ARL8- and kinesin-1-binding sites. This interaction inhibits coupling of lysosomes to kinesin-1 and, consequently, lysosome movement toward the cell periphery. We also find that ARL8 does not just recruit SKIP to the lysosomal membrane but also relieves SKIP autoinhibition, promoting kinesin-1-driven, anterograde lysosome transport. Finally, our analyses show that the largely disordered middle region of SKIP mediates self-association and that this self-association enhances the interaction of SKIP with kinesin-1. These findings indicate that SKIP is not just a passive connector of lysosome-bound ARL8 to kinesin-1 but is itself subject to intra- and inter-molecular interactions that regulate its function. We anticipate that similar organelle- or GTPase-induced conformational changes could regulate the activity of other kinesin adaptors.
细胞质中细胞器的长程运动依赖于与微管马达的偶联,这个过程通常由衔接蛋白介导。在许多情况下,这种偶联涉及细胞器或衔接蛋白通过自抑制状态的构象反转来激活微管马达。在此,我们表明,一种名为 SKIP(也称为 PLEKHM2)的衔接蛋白的类似调节机制起作用。SKIP 与溶酶体膜上的小 GTP 酶(GTPase)ARL8 结合,将溶酶体与正向微管马达 kinesin-1 偶联。SKIP 的结构-功能分析表明,包含三个 pleckstrin 同源(PH)结构域的 C 端区域与包含 ARL8 和 kinesin-1 结合位点的 N 端区域相互作用。这种相互作用抑制了溶酶体与 kinesin-1 的偶联,从而抑制了溶酶体向细胞边缘的运动。我们还发现,ARL8 不仅将 SKIP 招募到溶酶体膜上,而且还解除了 SKIP 的自抑制,促进了 kinesin-1 驱动的正向溶酶体运输。最后,我们的分析表明,SKIP 中大部分无序的中间区域介导了自身缔合,并且这种自缔合增强了 SKIP 与 kinesin-1 的相互作用。这些发现表明,SKIP 不仅是溶酶体结合的 ARL8 与 kinesin-1 的被动连接物,而且其本身受到分子内和分子间相互作用的调节,从而调节其功能。我们预计,类似的细胞器或 GTPase 诱导的构象变化可能会调节其他 kinesin 衔接蛋白的活性。