URPHYM-GEMO, The University of Namur, rue de Bruxelles, 61, Namur 5000 Belgium.
ncRNA, epigenetic and genome fluidity, Institut Curie, PSL Research University, CNRS UMR 3244, Université Pierre et Marie Curie, Paris, France.
Sci Adv. 2020 Dec 9;6(50). doi: 10.1126/sciadv.abc1450. Print 2020 Dec.
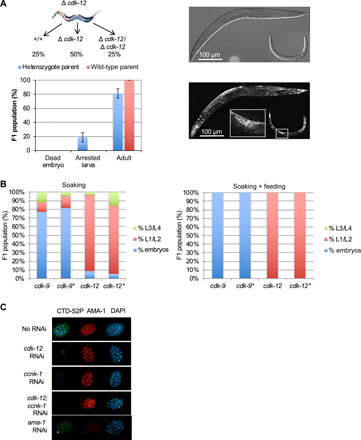
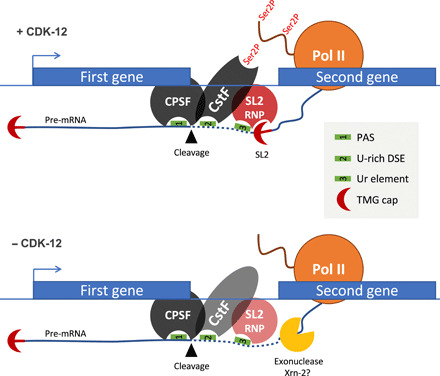
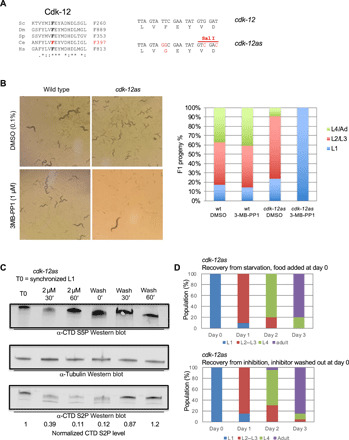
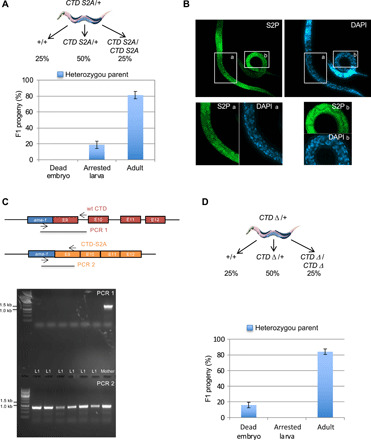
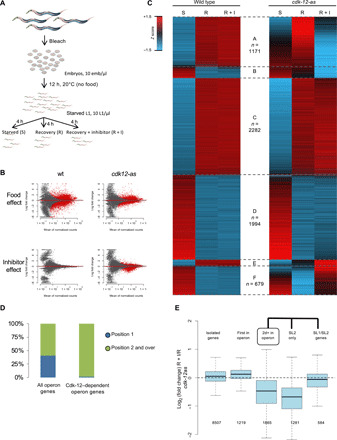
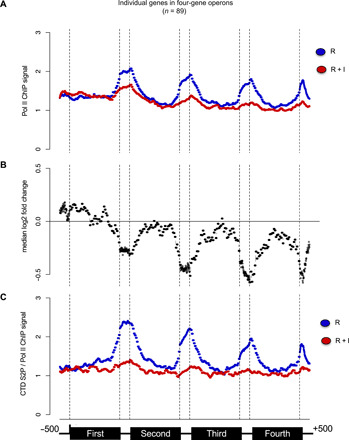
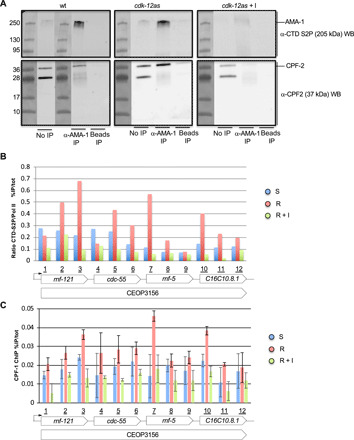
Serine 2 phosphorylation (S2P) within the CTD of RNA polymerase II is considered a Cdk9/Cdk12-dependent mark required for 3'-end processing. However, the relevance of CTD S2P in metazoan development is unknown. We show that lesions or a full-length CTD S2A substitution results in an identical phenotype in Embryogenesis occurs in the complete absence of S2P, but the hatched larvae arrest development, mimicking the diapause induced when hatching occurs in the absence of food. Genome-wide analyses indicate that when CTD S2P is inhibited, only a subset of growth-related genes is not properly expressed. These genes correspond to SL2 trans-spliced mRNAs located in position 2 and over within operons. We show that CDK-12 is required for maximal occupancy of cleavage stimulatory factor necessary for SL2 trans-splicing. We propose that CTD S2P functions as a gene-specific signaling mark ensuring the nutritional control of the developmental program.
丝氨酸 2 位磷酸化(S2P)是 RNA 聚合酶 II CTD 上的一个 Cdk9/Cdk12 依赖性标记,是 3'末端加工所必需的。然而,CTD S2P 在后生动物发育中的相关性尚不清楚。我们发现,缺失或全长 CTD S2A 取代会导致胚胎发生完全没有 S2P 的情况下产生相同的表型,但孵化的幼虫会停止发育,类似于在没有食物的情况下孵化时诱导的休眠。全基因组分析表明,当 CTD S2P 被抑制时,只有一部分与生长相关的基因不能正常表达。这些基因对应于位于操纵子 2 位和 2 位以上的 SL2 反式拼接 mRNA。我们表明 CDK-12 是最大程度占据切割刺激因子所必需的,而切割刺激因子是 SL2 反式拼接所必需的。我们提出,CTD S2P 作为一种基因特异性信号标记,确保了发育程序对营养的控制。